金桔
金币
威望
贡献
回帖0
精华
在线时间 小时
|
登陆有奖并可浏览互动!
您需要 登录 才可以下载或查看,没有账号?立即注册


×
第五篇 突触的相分离

https://doi.org/10.1038/s41593-019-0579-9
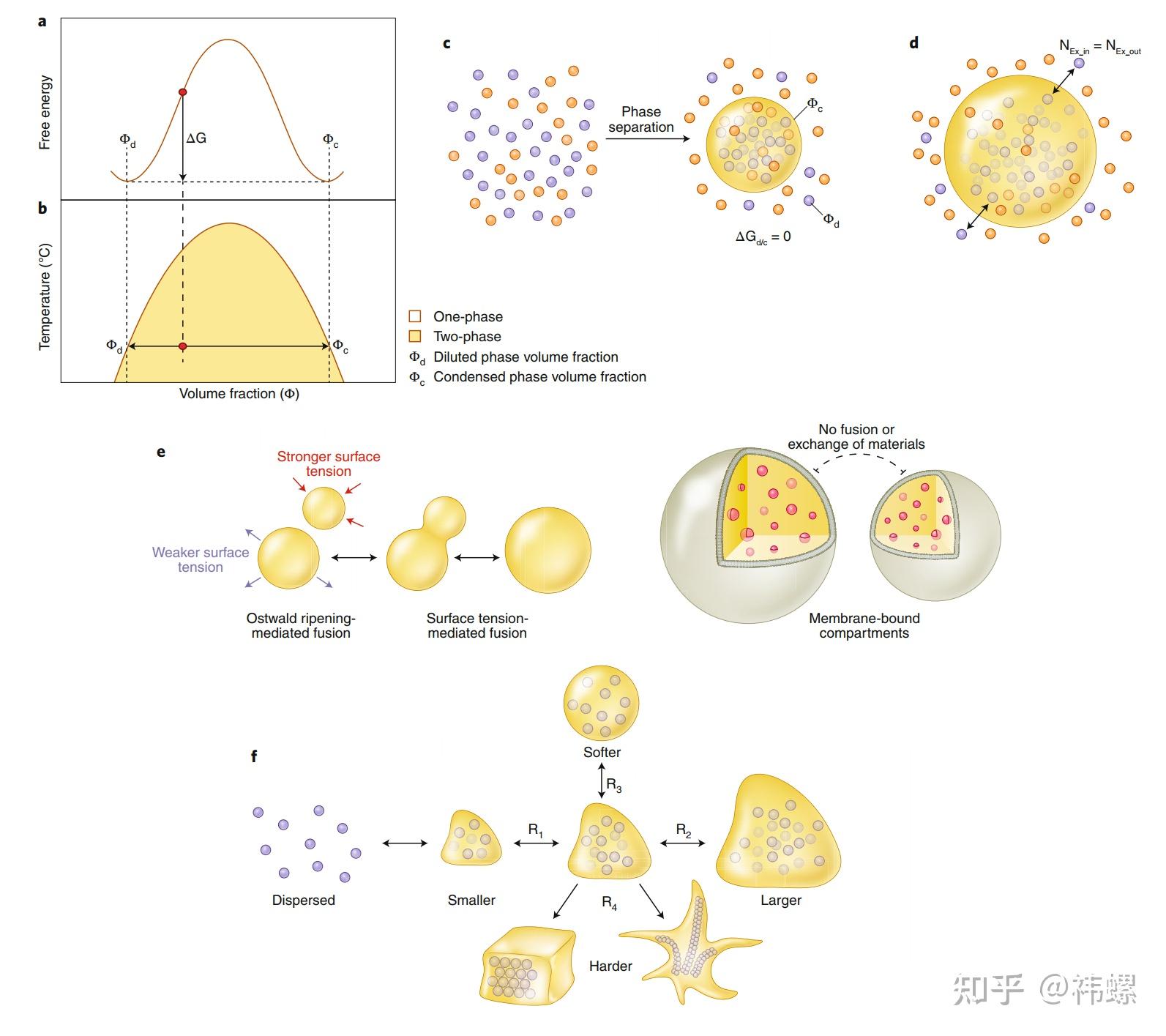
Basic principles of phase separation illustrated by a simple two-component system
a,自由能图显示了在一定条件下,两组分系统(例如,蓝色点表示的蛋白质在金色点表示的水溶液中)的相分离。一个均匀混合的系统可以通过降低自由能(G)达到最低水平而发生相分离,从而产生两相系统:一个稀溶相,其蛋白质浓度较低(0,以稀溶相的体积分数表示),和一个浓缩相,其蛋白质浓度较高(o,以浓缩相的体积分数表示)。
b,通过绘制自由能最小值作为温度的函数构建的两组分系统的相图。曲线表示系统从均匀的单相状态向两相状态转变的明显界限(或阈值浓度)。
c,相分离结果为浓缩相富集了特定的分子组分(这里以蓝色点表示),并被稀释相包围,稀释相中蓝点分子的浓度要低得多。
d,相分离后,两相之间的组分可以自由交换。然而,在两相之间没有组分的净流动,因为进出的分子数是相等的。浓缩相滴可以彼此融合成更大的滴,这种融合可以减小较小滴的表面张力。此外,小滴可以自发地收缩,释放的分子可以被其他更大的滴吸收,这不是一种材料交换的过程,而是一种称为奥斯特瓦尔德成熟的过程。
右侧是膜分隔的区域。生物凝聚体具有高度可塑性,并可以根据调节输入(R)经历不同形式的变化。通过简单地增加或减少系统中物质的数量(R和R2),可以使滴大小缩小或增大。通过不同的调节修改(R3和R4,例如形成可逆的凝胶状蛋白质聚集体或甚至一些不可逆的固体状蛋白质沉积物),滴可以变得更软或更硬。
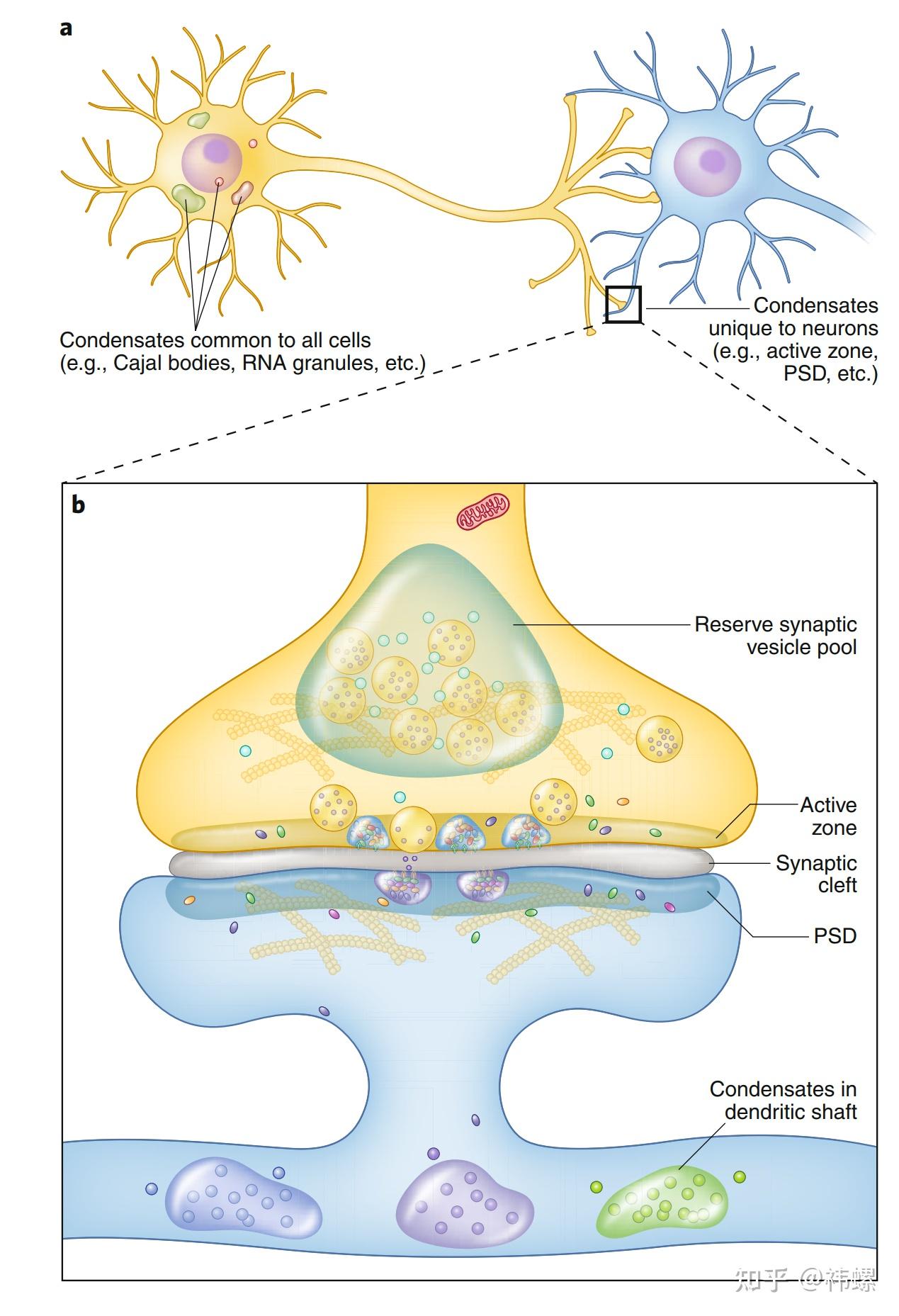
Phase separation in neurons.
a示意图展示了两个高度极化的神经元通过突触相互通信。神经元还包含其他细胞类型常见的各种生物凝聚体,包括细胞体内的RNA颗粒和应激颗粒,以及细胞核内的 Cajal 小体和核仁。此外,神经元还包含特有的生物凝聚体,如突触中特有的 PSD(Postsynaptic Density,突触后密度)和活性区域。
b图总览了突触中各种生物凝聚体。在突触前小结(presynaptic bouton)中,储备的突泡(SVs)可以通过蛋白质如 Synapsin 通过相分离而成团聚。形成突触前活性区域和 PSD 组装也可能涉及相分离。突触间隙(synaptic cleft)可能也包含浓缩的蛋白质组装。此外,树突干(dendritic shafts)中可能含有各种生物凝聚体,如RNA颗粒,用于局部蛋白质合成。
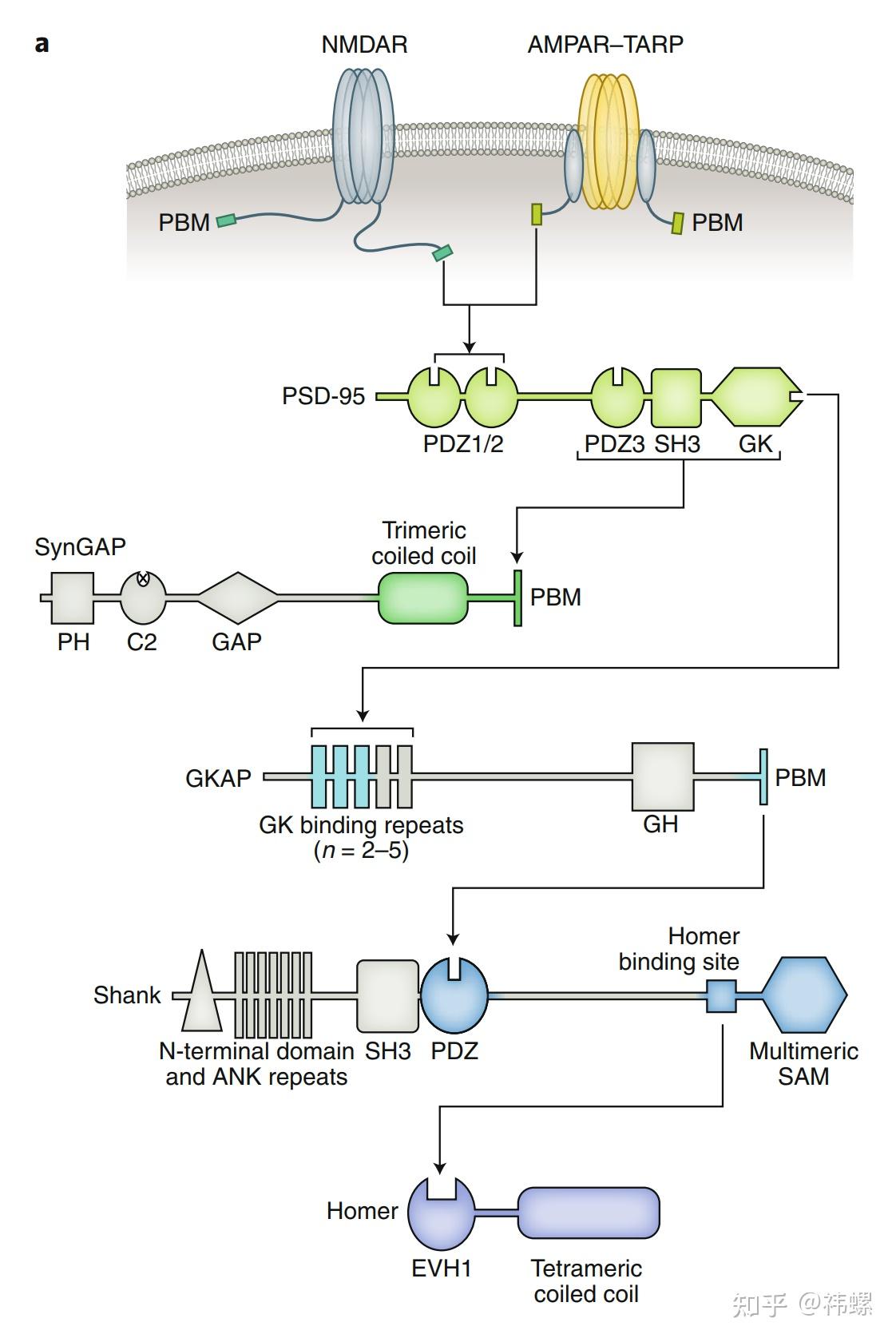
Schematic diagram showing the domain organizations and interaction networks ofkey PSD components.
示意图显示了关键 PSD 组分的域结构和相互作用网络。这些 PSD 组分包含重复的域或用于蛋白质相互作用或自身寡聚化的结构域,以增加网络的连通性。PSD 网络是由多价、特定、强烈的蛋白质相互作用形成的,可以在体外的生理蛋白质浓度下发生相分离。
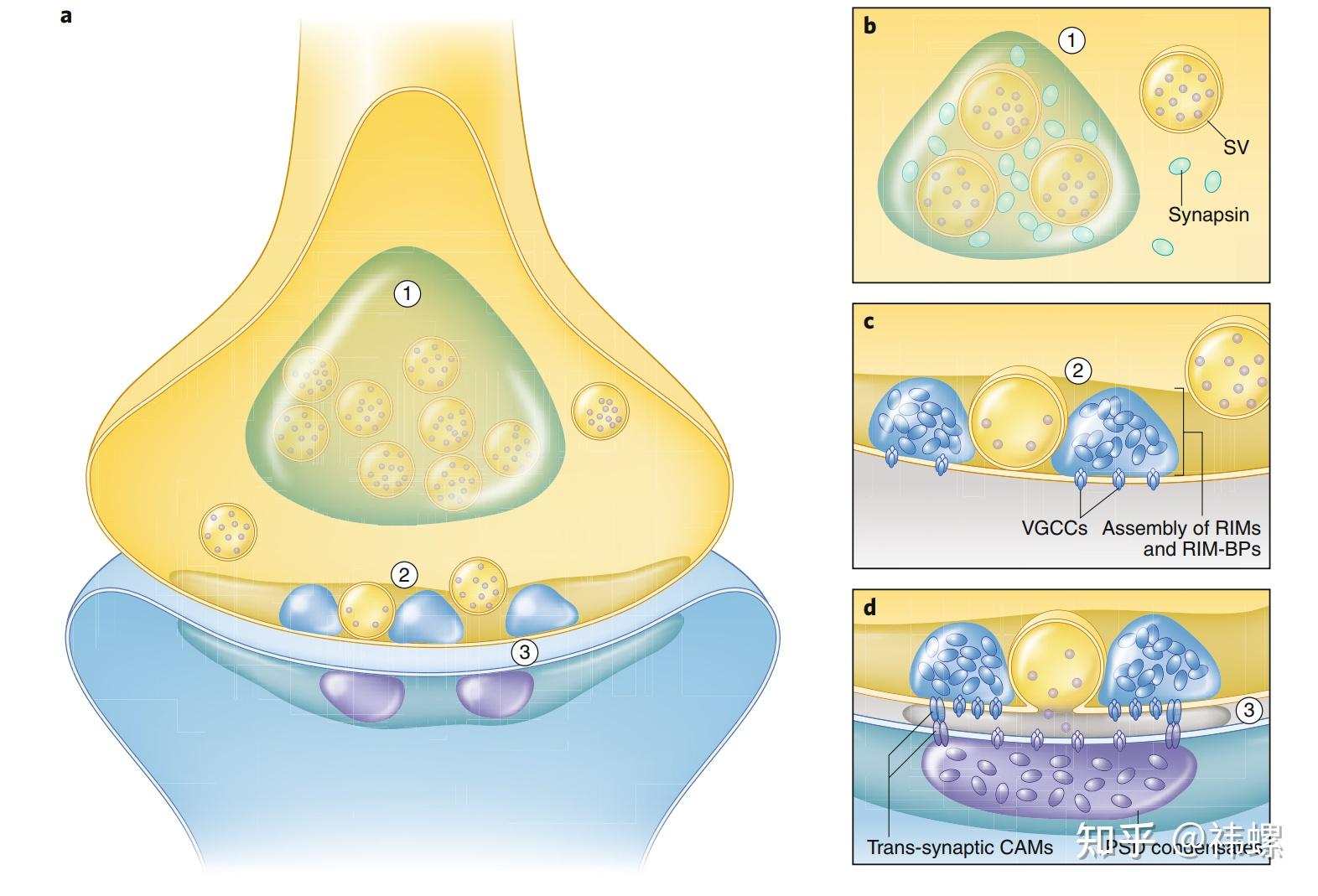
Phase separation in presynaptic boutons.
突触前小结中的相分离。a图是典型的兴奋性中枢神经系统突触中储备池 SV 相、活性区相和 PSD 相的分层排列概览。b图是通过 Synapsin 的相分离介导的储备池 SV 相形成以及 Synapsin 与 SVs 之间的相互作用的放大视图。c图显示了活性区密集投影凝聚体的形成,以及由 RIMs 和 RIM-BPs(以及可能其他突触前蛋白质如 ELKs、Munc13、Liprins 等)介导的 Ca2+ 通道聚集。图中还显示了密集投影凝聚体与停泊的 SVs 之间可能通过某些尚不清楚的机制进行直接相互作用。d图显示了突触前活性区凝聚体与 PSD 凝聚体的跨突触对齐,可能是通过跨突触细胞粘附分子(CAMs)实现的。
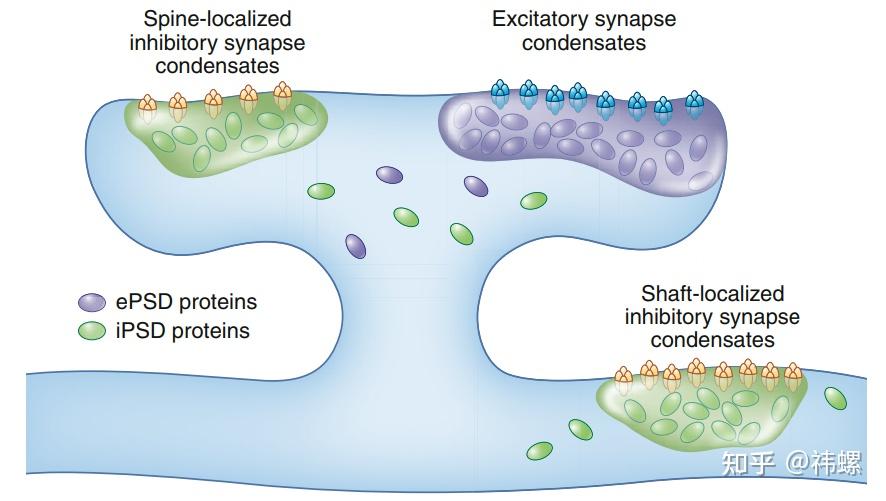
Mutual exclusion of excitatory and inhibitory PSD condensates.
兴奋性和抑制性PSD凝聚体相互排斥。图中示意模型显示了通过相分离介导的在微小的突刺头中分隔兴奋性和抑制性 PSD(分别为 ePSD 和 iPSD)凝聚体的情况。大多数抑制性 PSD 位于树突干上,因此自然地与位于树突刺上的兴奋性 PSD 分开。
<hr/>第六篇 转录控制的相分离模型

http://dx.doi.org/10.1016/j.cell.2017.02.007
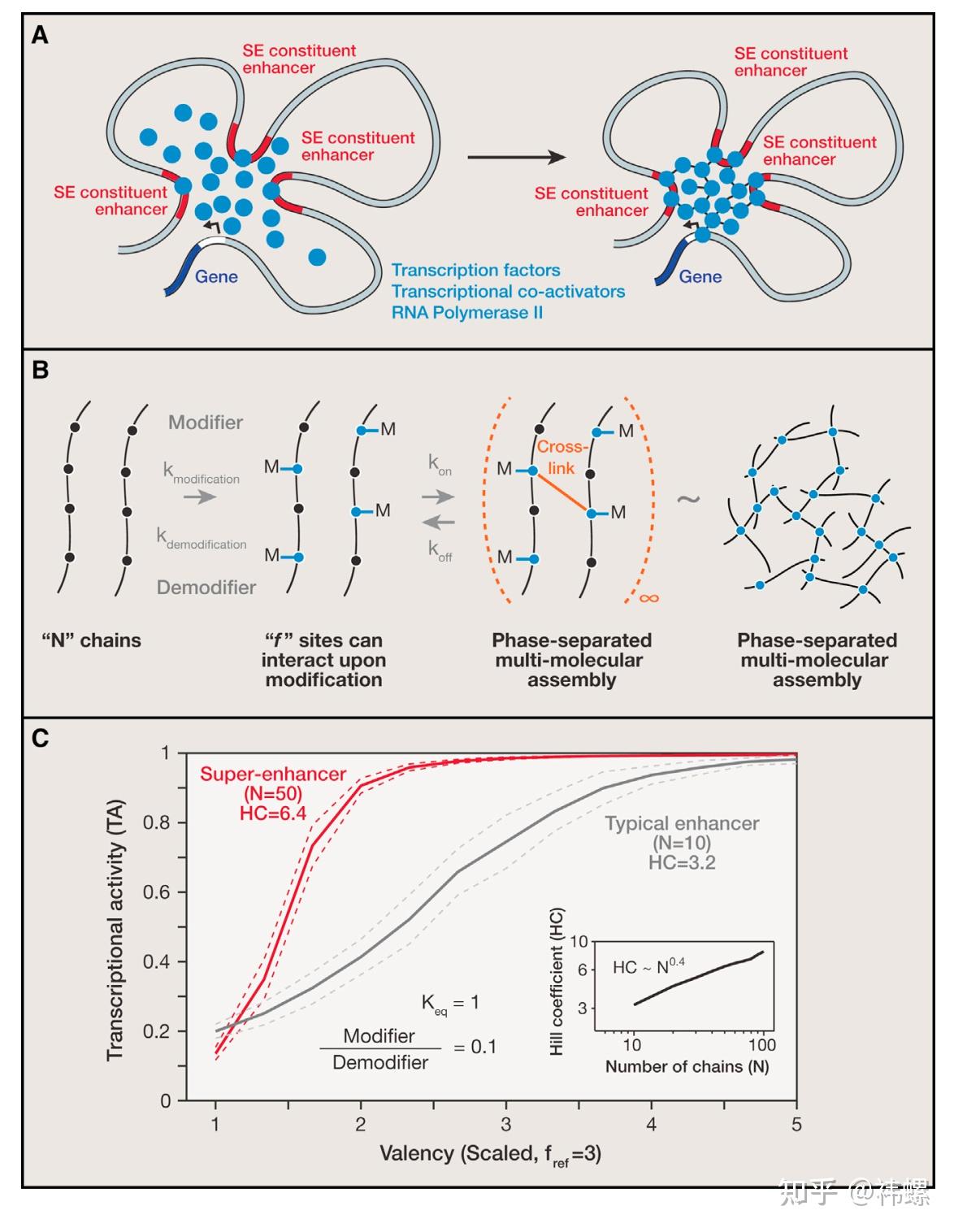
A Simple Phase Separation Modelof Transcriptional Control
(A) 生物系统的示意图,该系统可以在超增强子 - 基因座上形成相分离的多分子复合物,其中包括转录调控因子。
(B) 生物系统的简化表示,以及可能导致相分离的模型参数。其中“M”表示能够在修饰时形成交联的残基。
(C) 转录活性(TA)对超增强子(由 N = 50 链组成)和典型增强子(由 N = 10 链组成)的价数参数的依赖关系。代表转录活性(TA)的代理值被定义为交联链的最大簇的大小,按总链数进行缩放。价数经过缩放,以便实际价数除以一个参考数值为三。实线表示均值,虚线表示在50次模拟中的两倍标准偏差。K_eq 表示平衡常数,修饰物/去修饰物比例保持不变。HC 表示希尔系数,这是描述协同行为的经典度量标准。插图显示了希尔系数与系统中的链或组分数量的依赖关系。
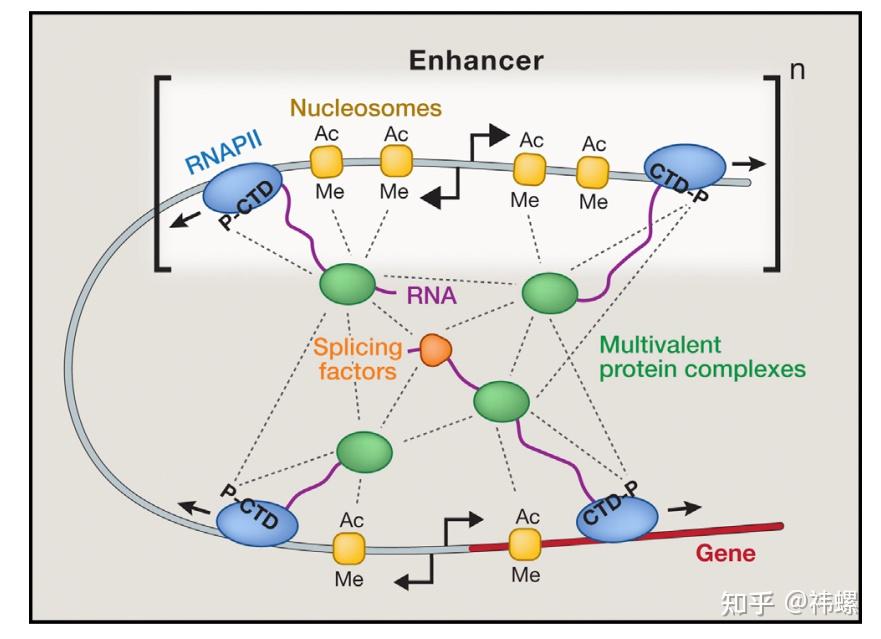
Transcriptional Control Phase Separation In Vivo
活体内的转录调控分相分离。基因调控元件中的相分离复合体模型。突出显示了形成复合体的一些候选转录调控因子。P-CTD 表示 RNA Pol II 的C-末端磷酸化区域。核小体的化学修饰(醋酸化,Ac;甲基化,Me)也被突出显示。增强子和启动子处的分散转录产生了可以被 RNA 剪接因子结合的初级 RNA。组件之间的潜在相互作用显示为虚线。
<hr/>第七篇 理解分相分离和解决未解问题

https://doi.org/10.1016/j.molcel.2022.05.018
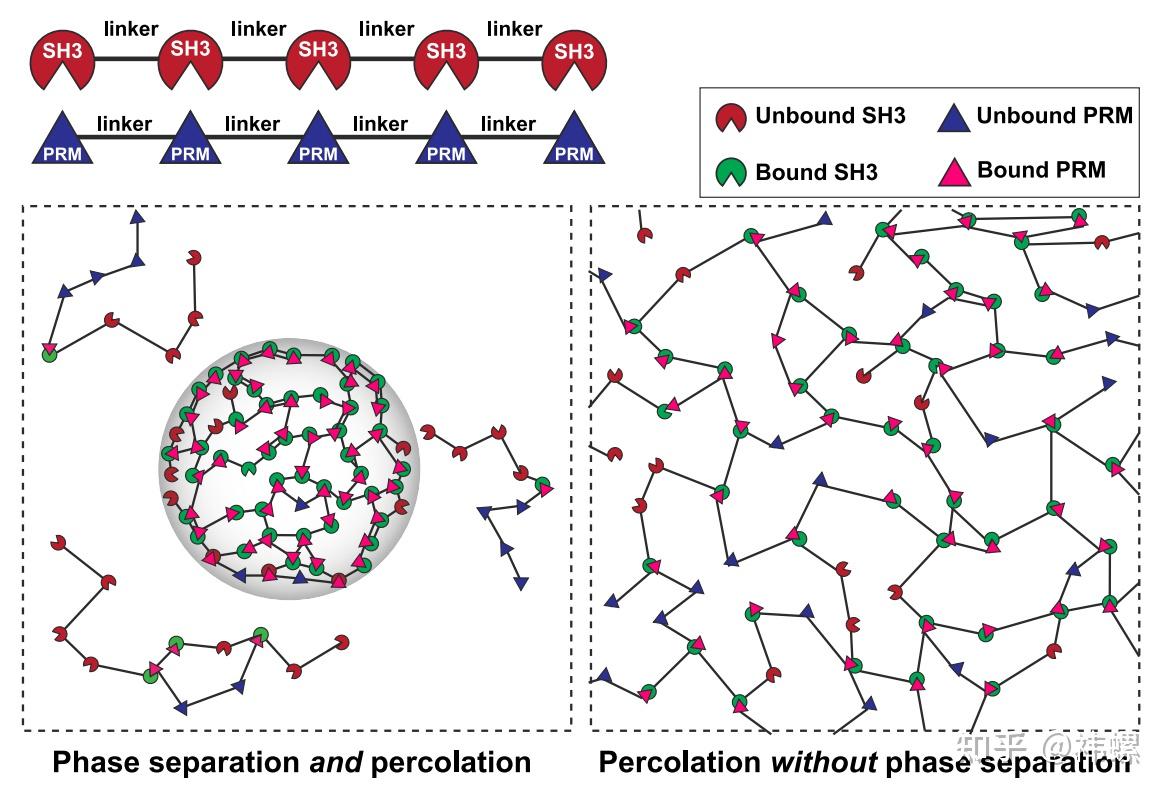
Coupling and decoupling of phaseseparation and percolation in multivalentproteins
两种蛋白质通过相互作用结构域和基序的重复进行相互作用,例如通过蛋白质 A 中 SH3 结构域的重复和蛋白质 B 中丰富的脯氨酸基序(PRMs)的重复(顶部);SH3 结构域和 PRMs 上的热点被称为“粘合点”。
多价相互作用介导了在渗透阈值 cperc 之上的渗透。单个蛋白质分子和复合物的溶解度,受连接子组成的强烈影响,决定了渗透是否与相分离相结合(如果csat < cperc,位于左侧),或者在没有相分离的情况下进行(如果cperc < csat,位于右侧)。
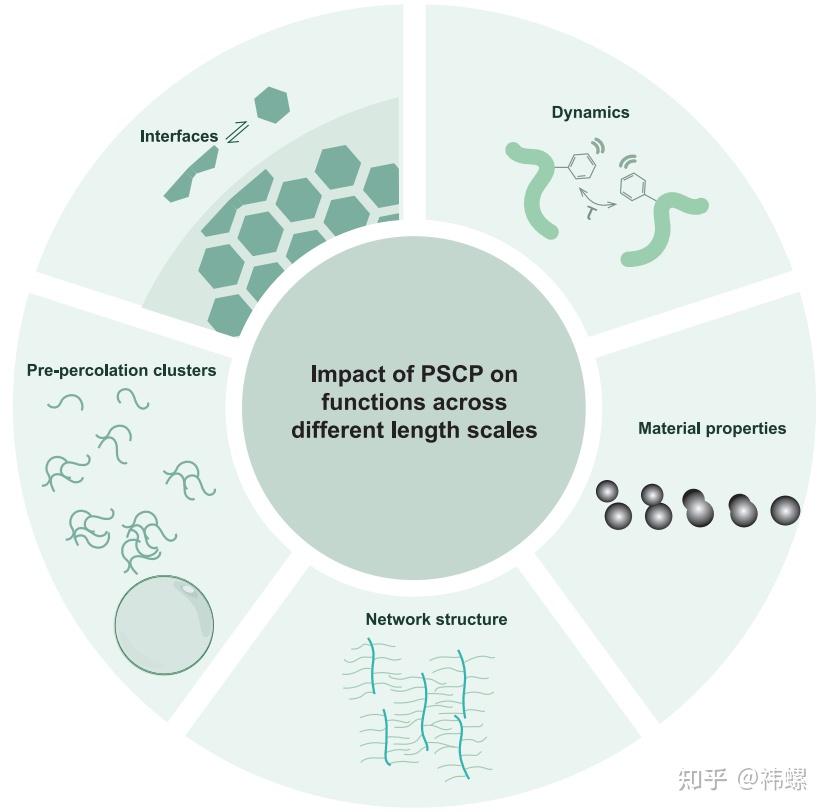
Consequences of condensates being viscoelasticnetwork fluids
通过相分离引起的多价大分子的性质决定了网络结构和动态、材料性质、界面性质以及渗透前团簇的大小分布。
<hr/>第八篇 相分离在聚合调控中的应用

https://doi.org/10.1016/j.jmb.2019.06.026
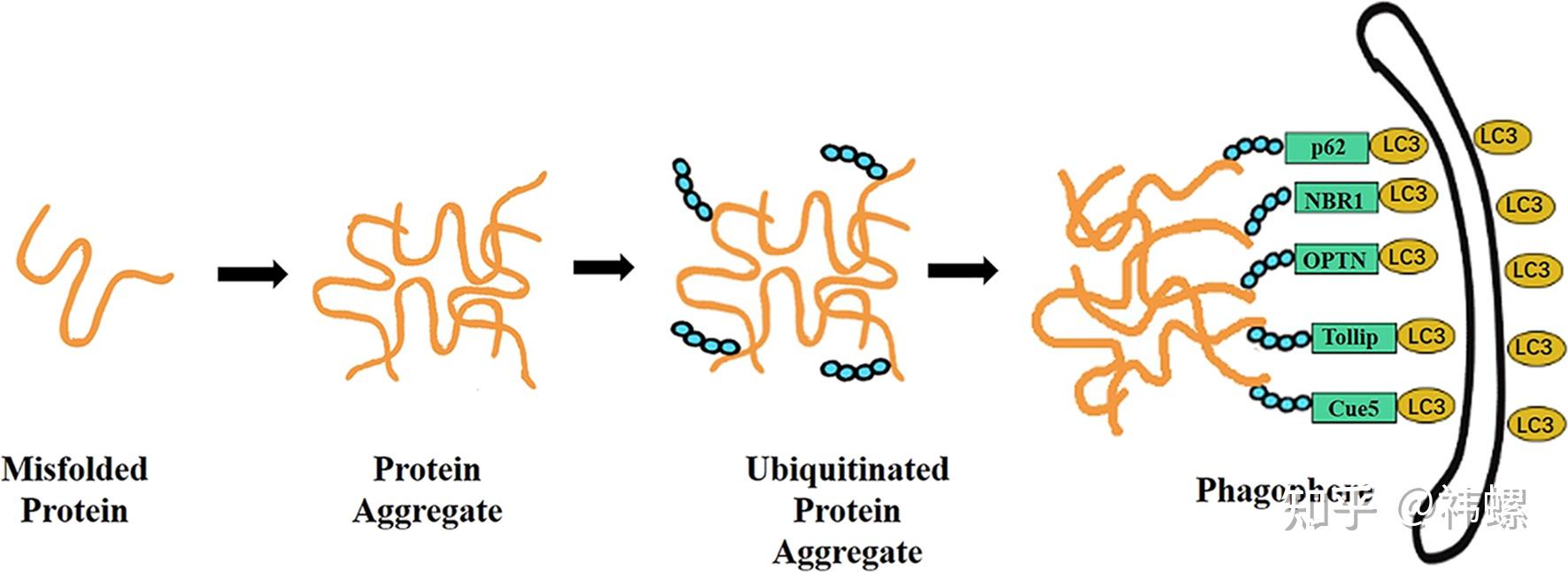
Aggrephagy.
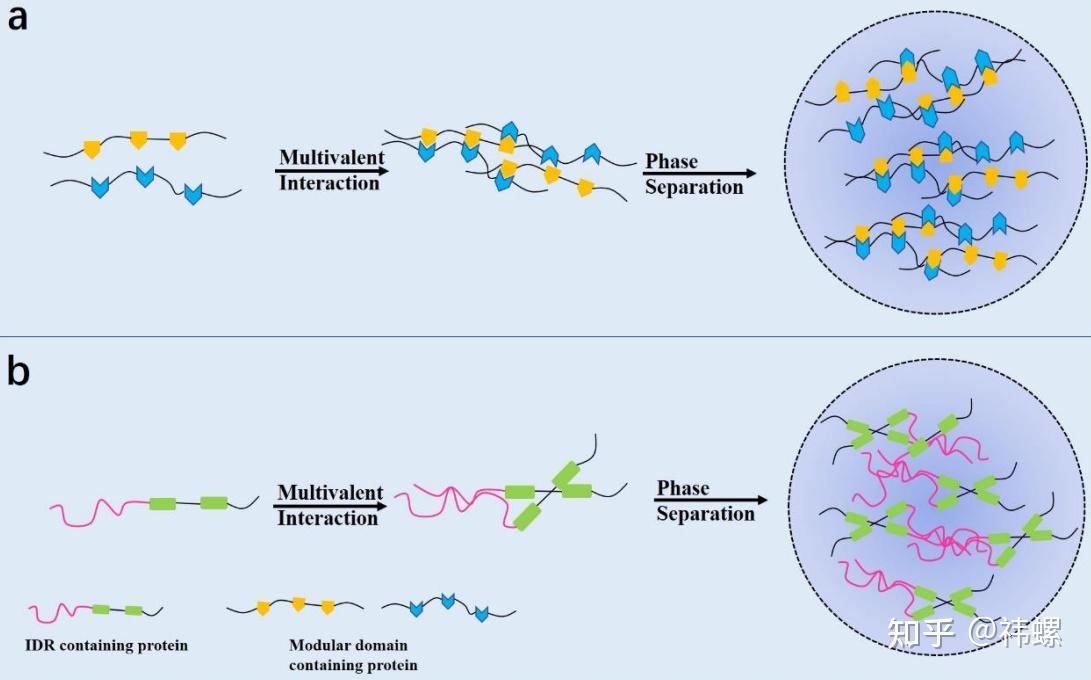
Multivalent interactions drive liquid-liquid phase separation in biological systems.
具有进行相分离能力的蛋白质通常拥有多个模块,可以创建多价性,包括模块化结构域(a)或本质上无序区域(IDRs)(b)。当蛋白质浓度达到临界阈值时,液-液相分离发生,形成无膜区。
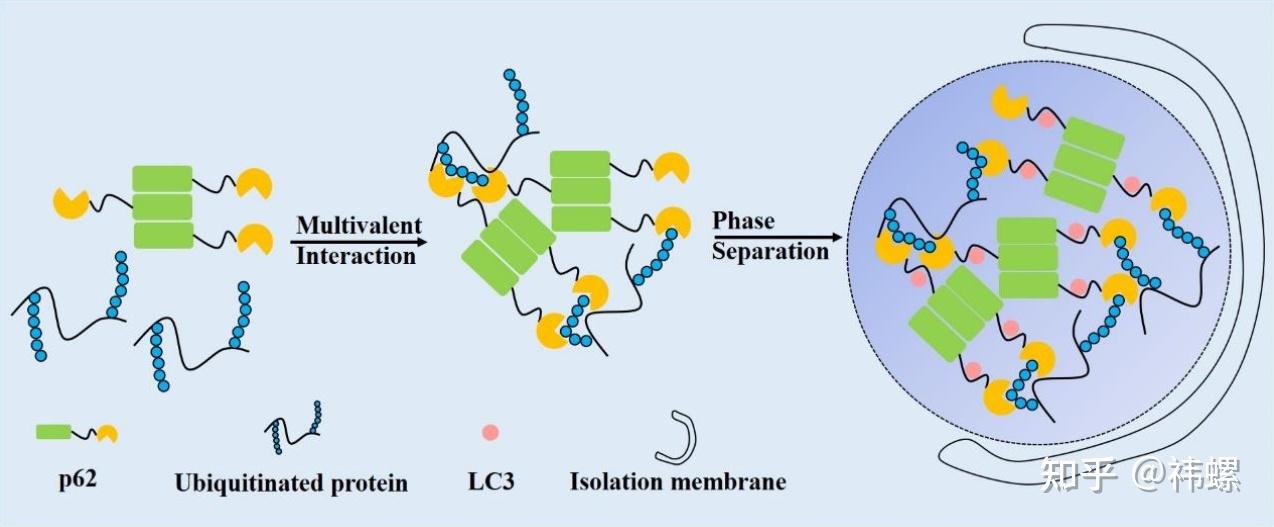
Polyubiquitin chain-induced p62 phase separation drives autophagic cargosegregation.
多泛泛素链诱导的 p62 相分离推动自噬的载体分离。p62 蛋白质通过 PB1 结构域(绿色部分)形成寡聚体,并通过 UBA 结构域(黄色部分)与泛泛素结合。这两个结构域促进多价相互作用。当蛋白质浓度达到阈值时,液-液相分离发生,形成 p62 体。其他客户蛋白,如 LC3 和 Keap1,也被招募到 p62 体中。然后,p62 体被自噬降解。
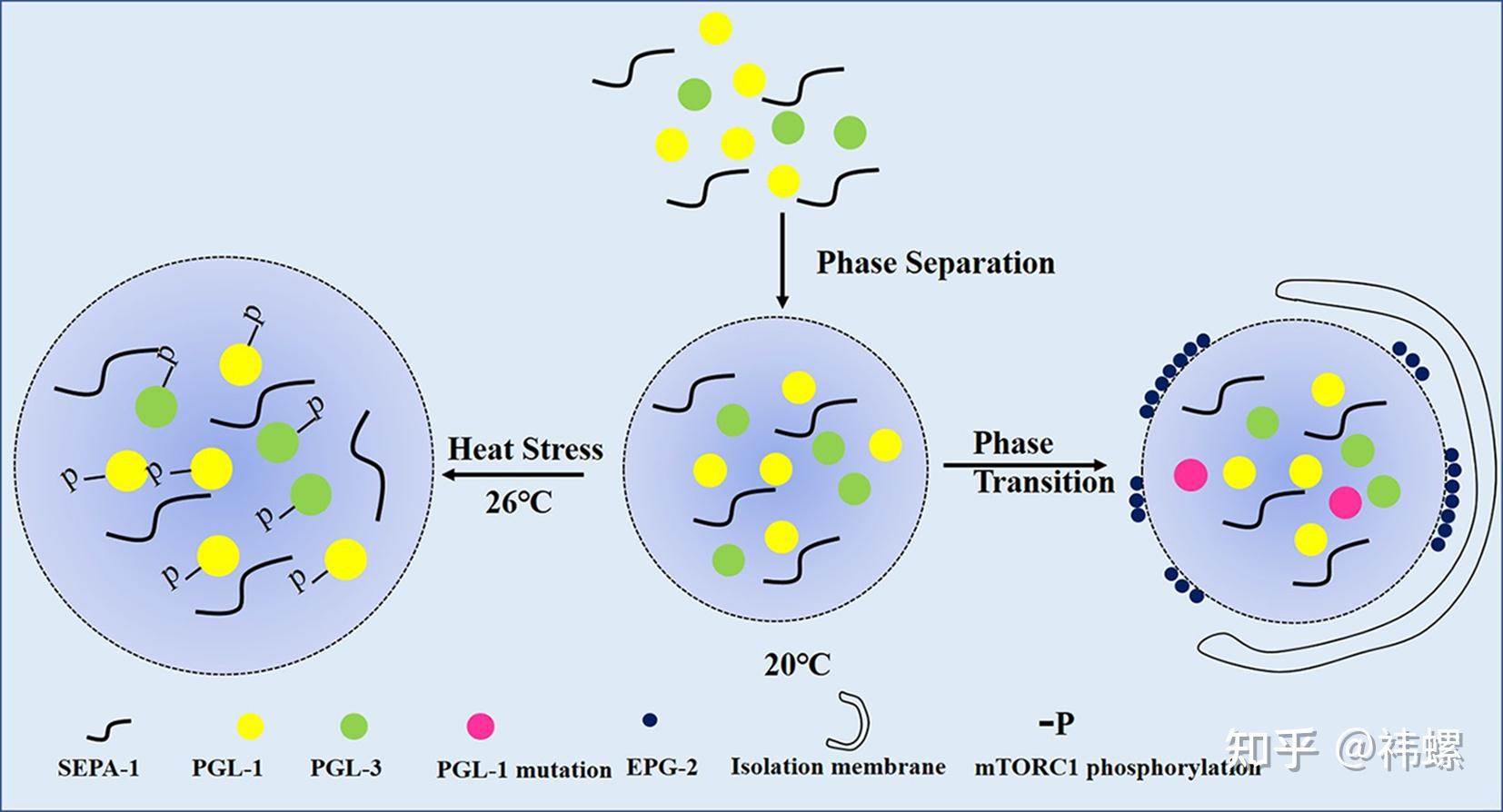
PGL granules are assembled via liquid-liquid phase separation and are degradedthrough autophagy.
PGL 颗粒是通过液-液相分离组装的,然后通过自噬降解。PGL-1 和 PGL-3 经历液-液相分离形成 PGL 颗粒。PGL-1/3 的相分离受 SEPA-1 的促进。EPG-2 和 PGL-1 的特定突变体(P55S、L82P、E360K)促进了 PGL 颗粒向凝胶状结构的相变,这些结构远不如以前那么流动,因此通过自噬降解。相反,在 26 °C 下,mTORC 对 PGL-1/3 的增强磷酸化促进了 PGL-1/3 的相分离,形成超过自噬降解的最佳尺寸的 PGL 颗粒。
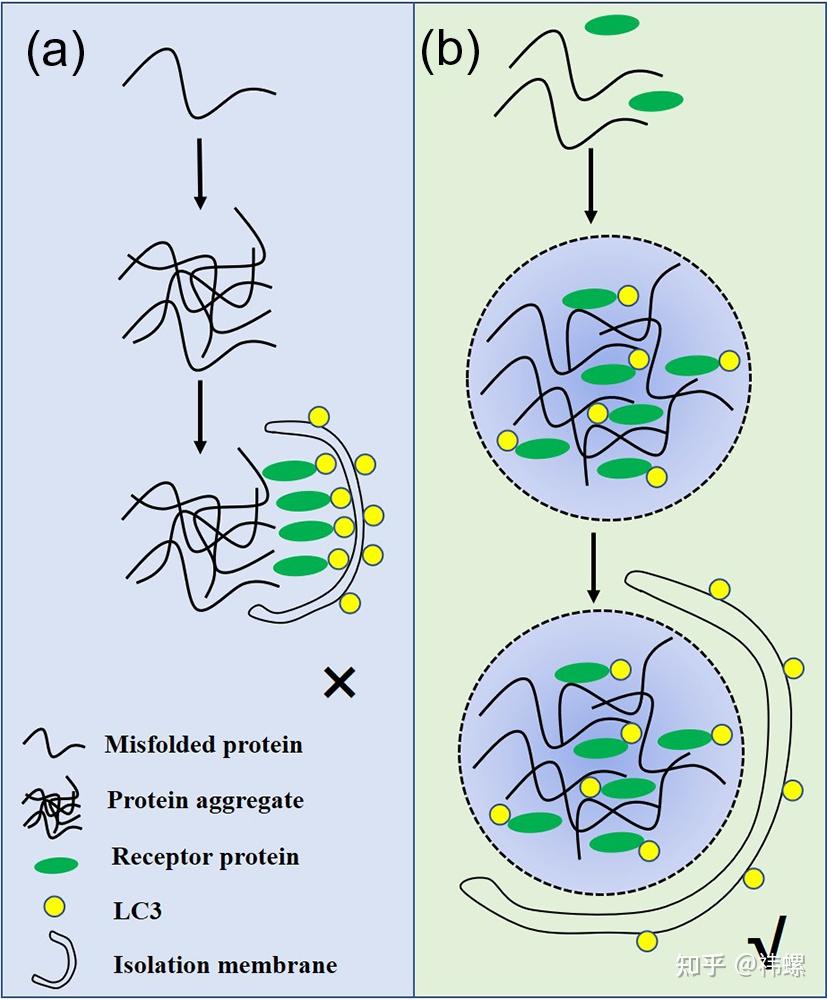
Aggrephagy cargos are segregated by liquid-liquid phase separation.
聚集体酶促自噬的载体通过液-液相分离进行分离。聚集体酶促自噬的载体通过液-液相分离形成类似液体的蛋白质液滴(b),而不是以前认为的简单蛋白质聚合物(a)。这些液滴的类似液体属性可能与它们被自噬机制识别相关。
<hr/>私信可获取文章下载链接
参考文献:
1. Hnisz D, Shrinivas K, Young RA, Chakraborty AK, Sharp PA. A Phase Separation Model for Transcriptional Control. Cell. 2017;169(1):13-23. doi:10.1016/j.cell.2017.02.007
2. Mittag T, Pappu R V. A conceptual framework for understanding phase separation and addressing open questions and challenges. Mol Cell. 2022;82(12):2201-2214. doi:10.1016/j.molcel.2022.05.018
3. Sun D, Wu R, Li P, Yu L. Phase Separation in Regulation of Aggrephagy. J Mol Biol. 2020;432(1):160-169. doi:10.1016/j.jmb.2019.06.026
4. Chen X, Wu X, Wu H, Zhang M. Phase separation at the synapse. Nat Neurosci. 2020;23(3):301-310. doi:10.1038/s41593-019-0579-9
原文地址:https://zhuanlan.zhihu.com/p/663015386 |
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 





 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 雷达卡
雷达卡 发表于 2025-2-19 21:27
发表于 2025-2-19 21:27
 提升卡
提升卡


