金桔
金币
威望
贡献
回帖0
精华
在线时间 小时
|
出分啦,87,祝大家都能取得满意的成绩!
<hr/>C值、蛋白质折叠非重点
A/B卷的出法→找那种对立可以分开考的知识点
同尾酶和同裂酶
克隆载体,表达
<hr/>考点
第二章
染色体具备哪些作为遗传物质的特征
DNA的一二三级结构
什么是DNA的tm值,受什么因素影响
什么是转座子,可分为哪些种类
什么是SNP?它作为第三代遗传标记有什么优点
第三章
大肠杆菌的RNA聚合酶有哪些组成成分?各个亚基的作用?
pribnow区
简述Ⅰ、Ⅱ类内含子的剪接特点
核酶具有哪些结构特点?核酶的生物学意义
三种剪接方式
mRNA的剪接
第四章
核糖体主要有哪些活性中心?
链霉素为什么能抑制蛋白质合成
哪些抗生素只能特异性地作用于原核生物核糖体?哪些只作用于真核生物核糖体?哪些能同时抑制原核,真核生物核糖体
什么是信号肽?它在序列组成上有哪些特点,有何功能
什么是核定位信号序列,其主要功能是什么
第五章
PCR技术
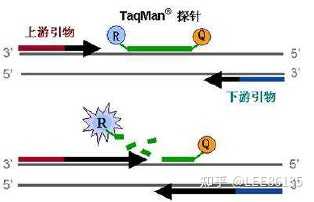
比较荧光染料sybrgreen Ⅰ和taqman荧光探针的主要不同点
分析基因组DNA文库和cDNA文库的主要区别
比较基因组与蛋白质组的主要差别
第六章
荧光原位杂交
RNAseq技术进行转录组学分析的原理
RNA原位杂交的主要实验过程及应用
在保持细胞及组织结构不变的条件下,用放射性或非放射性探针与组织切片反应,若细胞中存在能够特异性结合的mRNA序列,则产生双链RNA杂交体。通过放射性检验和酶促反应显色对基因的表达产物在细胞水平上做出定性、定量分析
检测动植物细胞中某种特定mRNA的分布状况和相对定量,进而进一步检测基因表达模式和基因表达量
免疫共沉淀实验的原理与过程,比较酵母双杂交技术和免疫共沉淀技术在研究蛋白质相互作用方面的优缺点
简述RNAi技术原理以及在分子生物学领域应用的前景
简述CRISPR/Cas9系统工作原理
简述酵母单,双杂交系统的基本原理与应用
凝胶滞缓实验的原理与应用
第七章
简述乳糖操纵子的调控模型
什么是葡萄糖效应
什么是弱化作用
应该不考
多启动子调控的操纵子、固氮基因调控
可能不考
魔斑
可能会考
转录水平上的其他调控方式、弱化子、操纵子
考
组成型剪接与选择性剪接、操纵子影响因子、葡萄糖对乳糖操纵子的影响、色氨酸操纵子
必考
重叠基因、翻译阻遏
名词解释
辅阻遏蛋白、基因、基因组、基因家族、基因簇
填空题
σ因子的调控作用--HSP、枯草芽孢杆菌、安慰诱导物是什么
选择题
IPTG、ONPG长什么样子
简答题
二组分调控系统
<hr/> 其他操纵子--间接考(存在性)
新冠病毒的发病机制(可能是给材料进行分析)
中和抗体--中和抗体是大分子药物,从新冠肺炎康复患者血清中分离获得,针对新冠病毒刺突蛋白受体结合域(RBD),包裹其侵入细胞的功能区域,阻碍病毒对细胞的感染,研发快,特异性强,副作用相对较小,疗效确切性强。北京大学生物医学前沿中心谢晓亮团队利用单细胞测序技术,取得重要进展。目前抗体DVP604经过大规模测试证明其活性、广谱性和抗病毒性均表现良好,且成本较低,可以作为新冠药物进入临床使用。
第八章
简述何为外显子,内含子及其结构特点和可变调控
简述DNA甲基化对基因表达的调控机制
简述真核生物转录元件组成及其分类
简述增强子的作用机制
可能考填空
反式作用因子、转录激活功能、反式作用因子能直接或间接地识别或结合在各类顺式作用元件的核心序列上
干湿实验
第九章
分析cart细胞疗法治疗癌症的优缺点
简述病毒载体与非病毒载体的优缺点
HBV病毒结构图考连线题
名词解释
病死率
群体免疫
可能会考
鸡尾酒疗法
第十章
哪些生物适合做模式生物
果蝇、斑马鱼、线虫、小鼠
第十一章
P435 人类基因组计划的8点科学意义
确定人体基因组中2万~2.5万个编码基因的序列及其在基因组中的物理位置
了解转录和剪接调控元件的结构与位置,从整个基因组的宏观水平上理解基因的转录与转录后调节
从整体上了解染色体结构,包括各种重复序列及非转录调控序列的大小和组织,了解不同序列在形成染色体结构,DNA复制,基因转录及表达调控中的影响与作用
简述新一代测序各平台的优点和缺点及适用领域
<hr/>
第一章 绪论
细胞学说要点
- 细胞是一个有机体,一切动植物都由细胞发育而来,并由细胞和细胞产物所构成
- 细胞是一个相对独立的单位,既有它自己的生命,又对与其它细胞共同组成的整体的生命起作用
- 新细胞可以从老细胞中产生
分子生物学简史
- 孕育阶段:1820年~1950年
- 创立阶段:1950年~1970年
- 发展阶段:1970年~至今
三条基本原理
- 构成生物体各类有机大分子的单体在不同生物中都是相同的
- 某一特定生物体所拥有的核酸和蛋白质分子决定了它的属性
- 生物体内一切有机大分子的建成都遵循共同的规则
现代分子生物学的研究内容
目的是将不同的DNA片段按照人们的设计定向连接起来,在特定的受体细胞中与载体同时复制并得到表达,产生影响受体细胞的新的遗传性状
可用来大量生产某些在正常细胞代谢中产量很低的多肽;定向改造某些生物的基因组结构,提高经济价值;进行基础研究
- 基因表达调控
- 生物大分子的结构功能
- 基因组、功能基因组与生物信息学
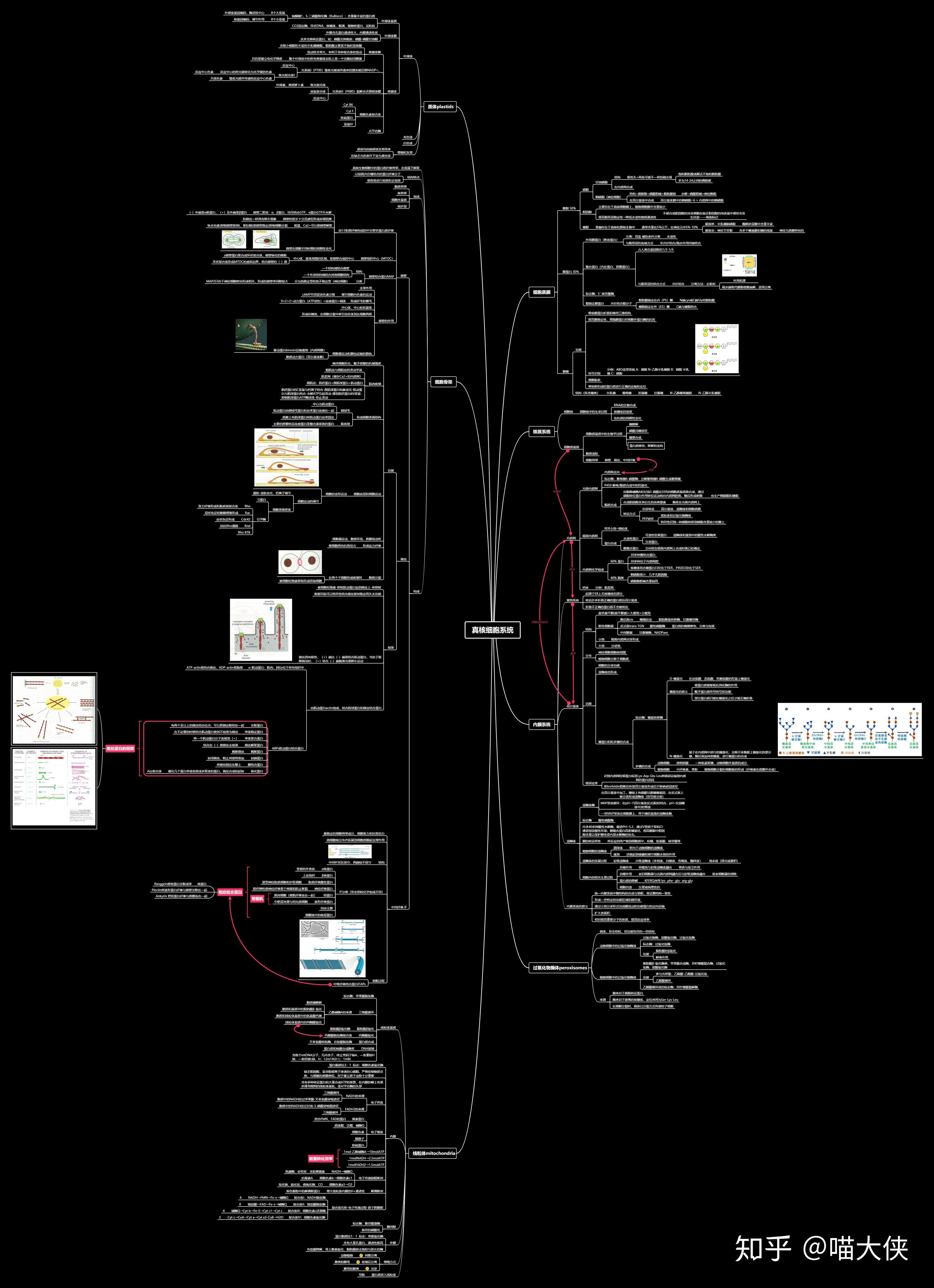
第二章 染色体与DNA
染色体(chromosome)
真核细胞染色体
- 本质是DNA和蛋白质的组合
- 是遗传信息的主要载体,但不是唯一载体
- 真核细胞位于细胞核的核仁内
染色体作为遗传物质的特征
- 分子相对稳定
- 能自我复制,保持遗传连续性
- 能指导蛋白质合成,控制生命过程
- 能产生可遗传的变异
蛋白质
组蛋白
是染色体的结构蛋白,它与DNA组成核小体。通常可以用2moI/L NaCI或0. 25moI/L的HCI/$$H_2SO_4$$,处理染色质,使组蛋白与DNA分开,然后用离子交换柱层析分离。
根据凝胶电泳性质,组蛋白分为H1、H2A、H2B、H3、H4。
这些组蛋白含有大量的赖氨酸和精氨酸,其中,H1富含赖氨酸,H3、H4富含精氨酸,H2A、H2B介于两者之间,呈碱性。
特性
- 进化上的极端保守性。牛,猪,大鼠的H氨基酸序列相同
- 无组织特异性
- 肽链上AA分布的不对称性。
- 组蛋白的修饰作用。包括甲基化,乙酰化,磷酸化,泛素化及ADP核糖基化等
- 富含赖氨酸的组蛋白H5
非组蛋白
高等动植物的核蛋白质中,除组蛋白之外的其他蛋白质的总称。呈酸性。
包括酶类(如RNA聚合酶),与细胞分裂有关的收缩蛋白,骨架蛋白,核孔复合物蛋白以及肌动蛋白,肌球蛋白,微管蛋白,原肌蛋白等
非组蛋白的量大约是组蛋白的60%~70%。有如下主要类型:
- HMG蛋白(high mobility group protein)
- DNA结合蛋白
- A24非组蛋白
C值--一种生物单倍体基因组DNA的总量
C值一般随生物进化而增加,但也存在某些低等生物的C值大于高等生物的现象
DNA
根据对DNA的动力学研究,真核细胞DNA序列分为
- 不重复序列:占40%~80%, 是主要的结构基因。如蛋清蛋白,蚕的丝心蛋白,血红蛋白都是单拷贝基因
- 中度重复序列:占10%~40%, 重复次数$$10^1\sim10^4$$, 包括各种rRNA和tRNA及结构基因
- 高度重复序列--卫星DNA,这类DNA只在真核生物中发现。可能与染色体稳定性有关
染色质和核小体
染色质由核小体组成,核小体由DNA与组蛋白八聚体构成
染色质DNA的Tm值比自由DNA高,说明在染色质中DNA极可能与蛋白质分子相互作用。在染色质状态下,由DNA聚合酶和RNA聚合酶催化的DNA复制和转录活性大大低于在自由DNA中的反应。
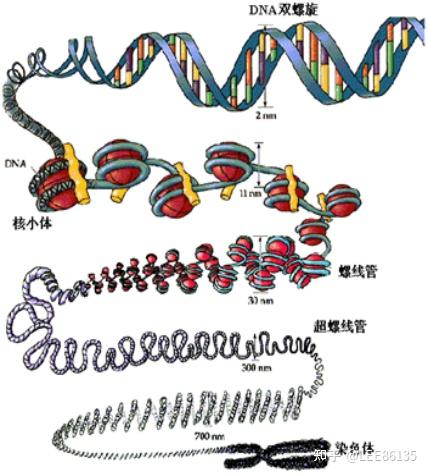
真核生物基因组特点
- 庞大
- 存在大量重复序列
- 大部分为非编码序列
- 转录产物为单顺反子
- 断裂基因,有内含子结构
- 存在大量顺式作用元件
- 存在大量的DNA多态性
- 存在端粒结构
原核生物基因组特点
- 主要为单拷贝基因,只有少数以多拷贝形式存在
- 几乎全部由功能基因和调控序列组成
- 几乎每个基因序列都与编码的蛋白质呈线性对应状态
<hr/>
基因组特点
DNA的结构
一级结构
4种脱氧核苷酸的排列
二级结构
两条核苷酸链反向平行缠绕所形成的双螺旋结构,分为右手螺旋和左手螺旋
高级结构--超螺旋结构
正超螺旋与负超螺旋
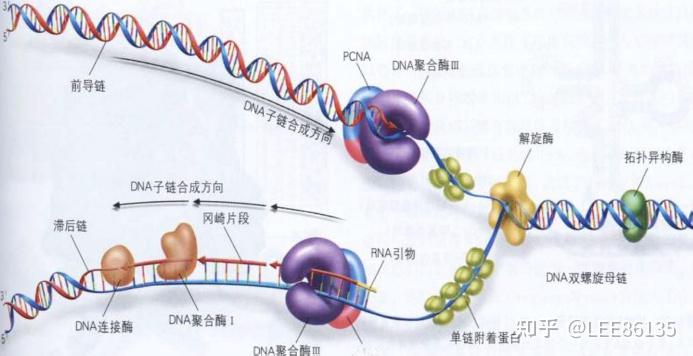
DNA的复制
半保留复制--DNA生物合成时,母链DNA解开为两股单链,各自作为模板按碱基配对规律,合成与模板互补的子链。子代细胞的DNA,一股单链从亲代完整地接受过来,另一股单链则完全从新合成。两个子细胞的DNA都和亲代DNA碱基序列一致。
半不连续复制--领头链连续复制而随从链不连续复制
双向复制、高保真性
复制子--两个相邻起始点之间的距离。复制子是独立完成复制的功能单位。
底物:dATP,dGTP,dCTP,dTTP
起点:复制叉
方向:5&#39;到3&#39;
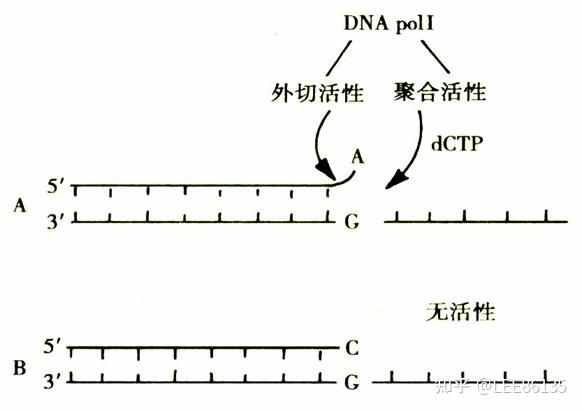
DNA聚合酶--依赖DNA的DNA聚合酶
- 5&#39;→3&#39;的聚合活性--能辨认错配的碱基对,并将其水解
- 核酸外切酶活性--能切除突变的DNA片段
原核生物
DNA polⅠ--对复制中的错误进行校读,对复制和修复中出现的空隙进行填补
DNA polⅡ--参与DNA损伤的应急状态修复;即使pol Ⅱ基因突变,细菌也能存活
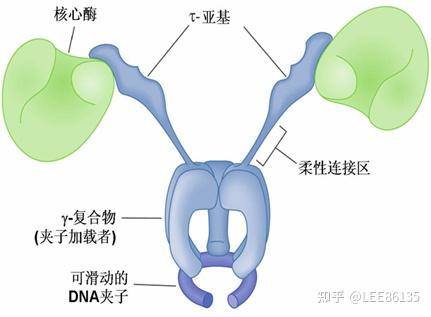
DNA polⅢ
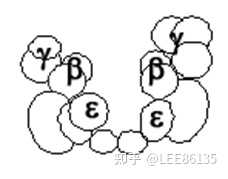
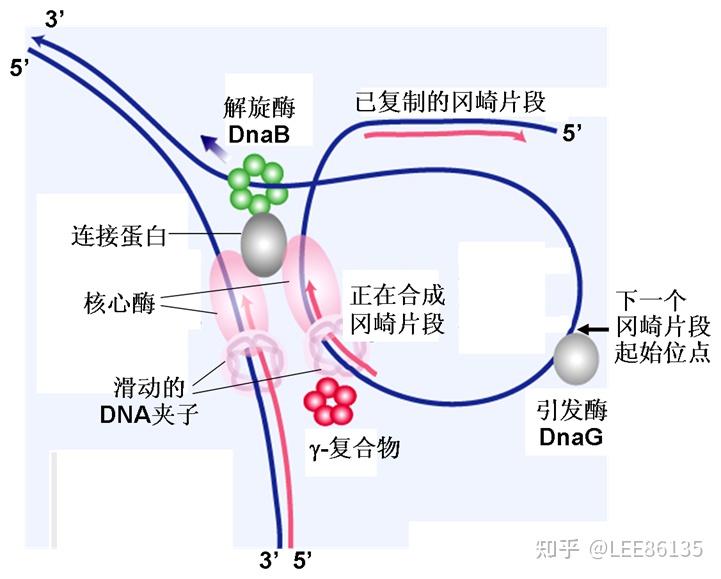
复制起始的相关蛋白质
解螺旋酶
利用ATP供能,作用于氢键,使DNA双链解开成为两条单链
拓扑异构酶
既能水解,又能连接磷酸二酯键
DNA连接酶
DNA连接酶接合复制、修复、重组及剪接中的缺口。也是基因工程的重要工具酶
真核细胞
DNA聚合酶
- DNAPα--起始引发,有引物酶活性
- DNAPβ--参与低保真度的复制
- DNAPγ--催化线粒体DNA复制
- DNAPδ--延长子链的主要酶,有解螺旋酶活性
- DNAPε--在复制中起校读、修复和填补缺口的作用
DNA复制的保真性
DNA聚合酶靠其大分子结构协调非共价(氢键)与共价(磷酸二酯键)键的有序形成。
嘌呤的化学结构能形成顺式和反式构型。与相应的嘧啶形成氢键配对,嘌呤应处于反式构型
- 聚合酶在复制延长时对碱基的选择功能
- 复制出错时有即时校对功能
DNA聚合酶如何知道从哪里开始复制?
DNA聚合酶靠合成的一段RNA单链作为引物,指导从哪开始复制。(PCNA是聚合酶的辅助蛋白)
引物--一段DNA或RNA单链。 通过碱基互补配对定位复制的起始位置,告诉聚合酶从哪里开始复制
引发体--解螺旋酶、DNA B、DNA C蛋白、引物酶和DNA复制起始区域的复合结构体
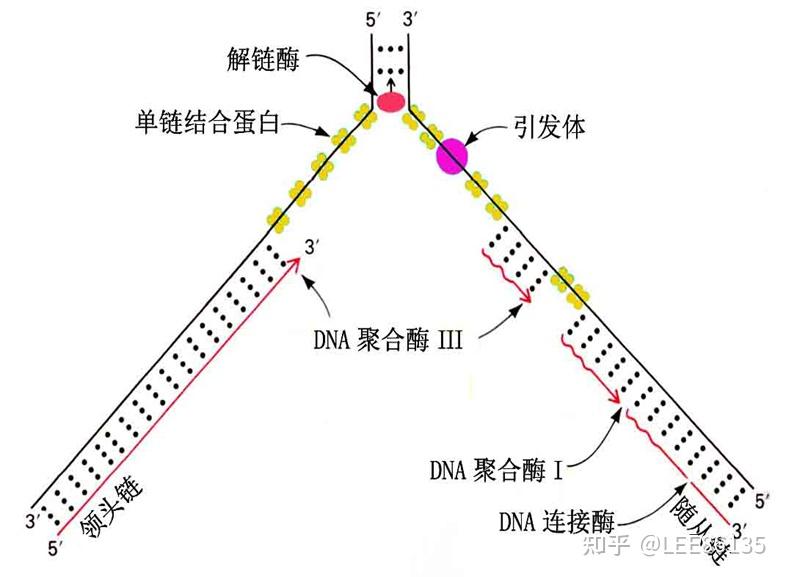
复制终止--切除RNA引物,填补空缺,连接切口
细胞能否分裂,决定于进入S期及M期这两个关键点。G1→S及G2→M的调节,与蛋白激酶活性有关。
蛋白激酶通过磷酸化激活或抑制各种复制因子而实施调控作用
增殖细胞核抗原(proliferation cell nuclear antigen,PCNA)在复制起始和延长中起关键作用。PCNA为同源三聚体,具有与E.coli DNA 聚合酶Ⅲ的β亚基相同的功能和相似的构象,即形成闭合环形的可滑动DNA夹子,在RFC的作用下PCNA结合于引物-模板链;并且PCNA使polδ获得持续合成能力。PCNA水平也是检验细胞增殖的重要指标
DNA-pol δ和pol α分别兼有解螺旋酶和引物酶活性。在复制叉及引物生成后,DNA-pol δ通过PCNA的协同作用,逐步取代pol α,需要拓扑酶和复制因子在RNA引物的3&#39;-OH基础上连续合成领头链。随从链引物也由pol α催化合成。然后由PCNA协同,pol δ置换pol α,继续合成DNA子链
真核生物DNA合成后立即组装成核小体
真核生物冈崎片段长度小于原核生物
真核生物每个染色体有多个起始点,复制有时序性,即复制子以分组方式激活而不是同步起动
端粒
端粒--真核生物染色体线性DNA分子末端的结构
结构特点
由末端单链DNA序列和蛋白质构成。
末端DNA序列是多次重复的富含TG碱基的短序列。
功能:维持染色体的稳定性;维持DNA复制的完整性
复制最后的RNA引物被去除后形成空隙,端粒减少
端粒酶由端粒酶RNA(hTR)、端粒酶协同蛋白(hTP1)、端粒酶逆转录酶(hTRT)组成
复制的几种主要方式
- 线性DNA双链的复制
- 环状双链DNA的复制( θ型--大肠杆菌、滚环型、D-环形型--哺乳动物线粒体DNA)
无法复制加载中的内容
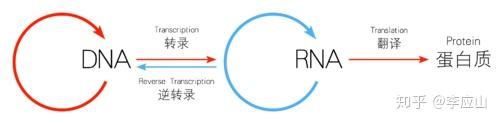
逆转录
RNA病毒在细胞内复制成双链DNA。这个DNA保留了RNA病毒全部遗传信息,并可在细胞内独立繁殖。在某些情况下,这个DNA通过基因重组,参加到细胞基因组内,并随宿主基因一起复制和表达。这种重组方式称为整合
cDNA--以mRNA为模板,经逆转录合成的与mRNA碱基序列互补的DNA链
cDNA法--应用逆转录酶获取基因工程目的基因
滚环复制
某些低等生物如M13噬菌体的复制方式
在复制过程中,DNA的双环沿着内环先滚一次,然后沿着外环再滚一次,内外环是不同时合成的,但是最终的结果和亲代完全一样的
D环复制
真核生物线粒体
在DNA-pol γ的催化下,从第一个起始点沿着内环合成新的子链,开环到一定的程度,在外环上的第二个起始点,又开始合成第二个引物。
D环复制有时间差,有独立的起始点

DNA的突变与修复
DNA突变/DNA损伤--个别dNTP残基或DNA片段在构成、复制或表型功能的异常变化
基因多态性一词用来描述个体之间的基因表型差别现象。
错配/点突变、缺失、插入、重排→框移突变→复制/转录障碍;DNA序列永久改变
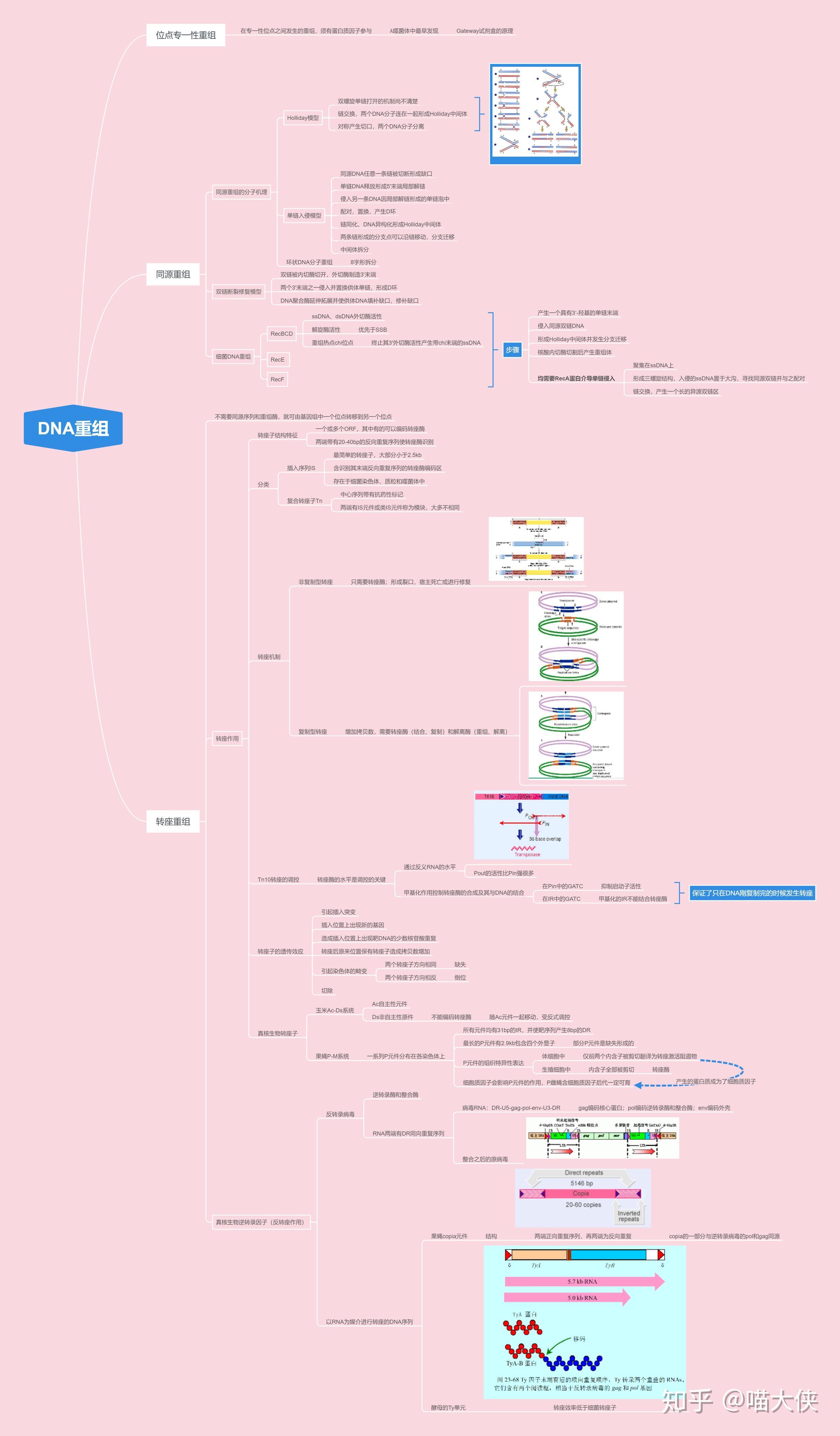
DNA的转座
由可移位因子介导的遗传物质重排现象
插入序列(IS因子)
- 含短的末端反向重复序列
- 含编码转座酶的基因
- 靶位点存在短正向重复序列
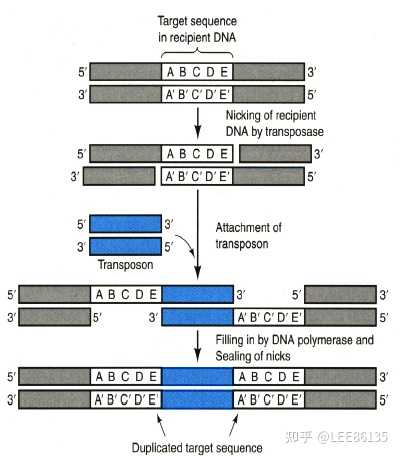
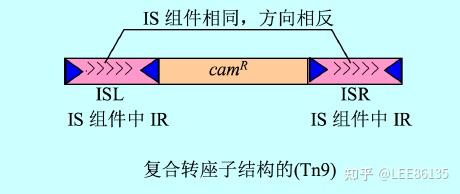
复合型转座子
- 有一类较大的可移动成分
- 除有关转座的基因外,至少带一个与转座作用无关并决定宿主菌遗传形状的基因(某些抗药性基因)
- 两翼往往为两个相同或高度同源的IS序列
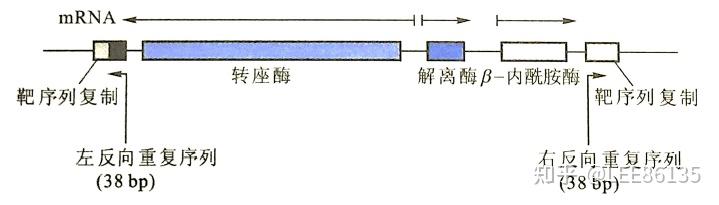
TnA家族
两端不含IS、带有转座酶基因、解离酶基因(调节基因)和抗生素基因
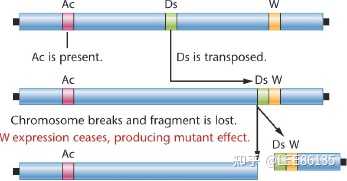
真核生物中的转座子
玉米
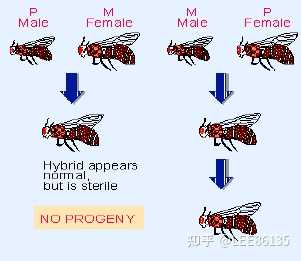
果蝇
转座作用机制
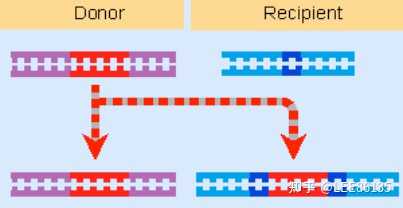

转座子元件被复制并移动到受体位点,最终转座过程扩增了转座子的拷贝(供、受点)
需两种酶:转座酶(作用于原拷贝两末端)、解离酶(作用于复制后的拷贝)
供体上最终产生双链断裂,阻止复制形成,不能形成共合体。供体位点如不能修复则有致死效应
转座作用的遗传学效应
- 转座引起突变
- 转座产生染色体畸变
- 转座引起新的基因
- 转座引起生物进化
SNP的理论与应用
传统检测方法(定性)--FRLP、PCR-SSCP、CE、DHPLC
国际通行检测方法(定量)--DNA测序
检测技术
- 基因芯片技术
- Taqman技术(供体-受体染料对,FRET)
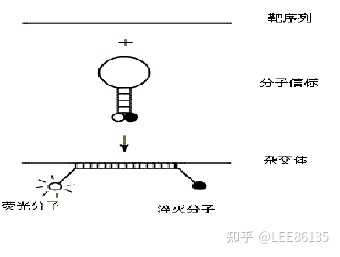
- 分子灯标技术
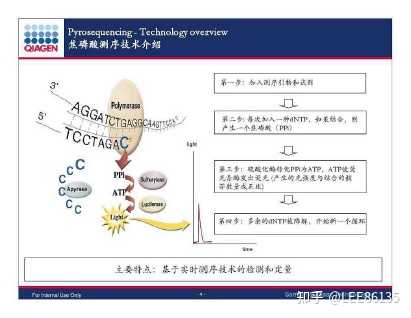
- 焦磷酸测序法
应用
- 人类单倍型图谱绘制
- SNP与疾病易感基因的相关性分析
- 指导用药与药物设计
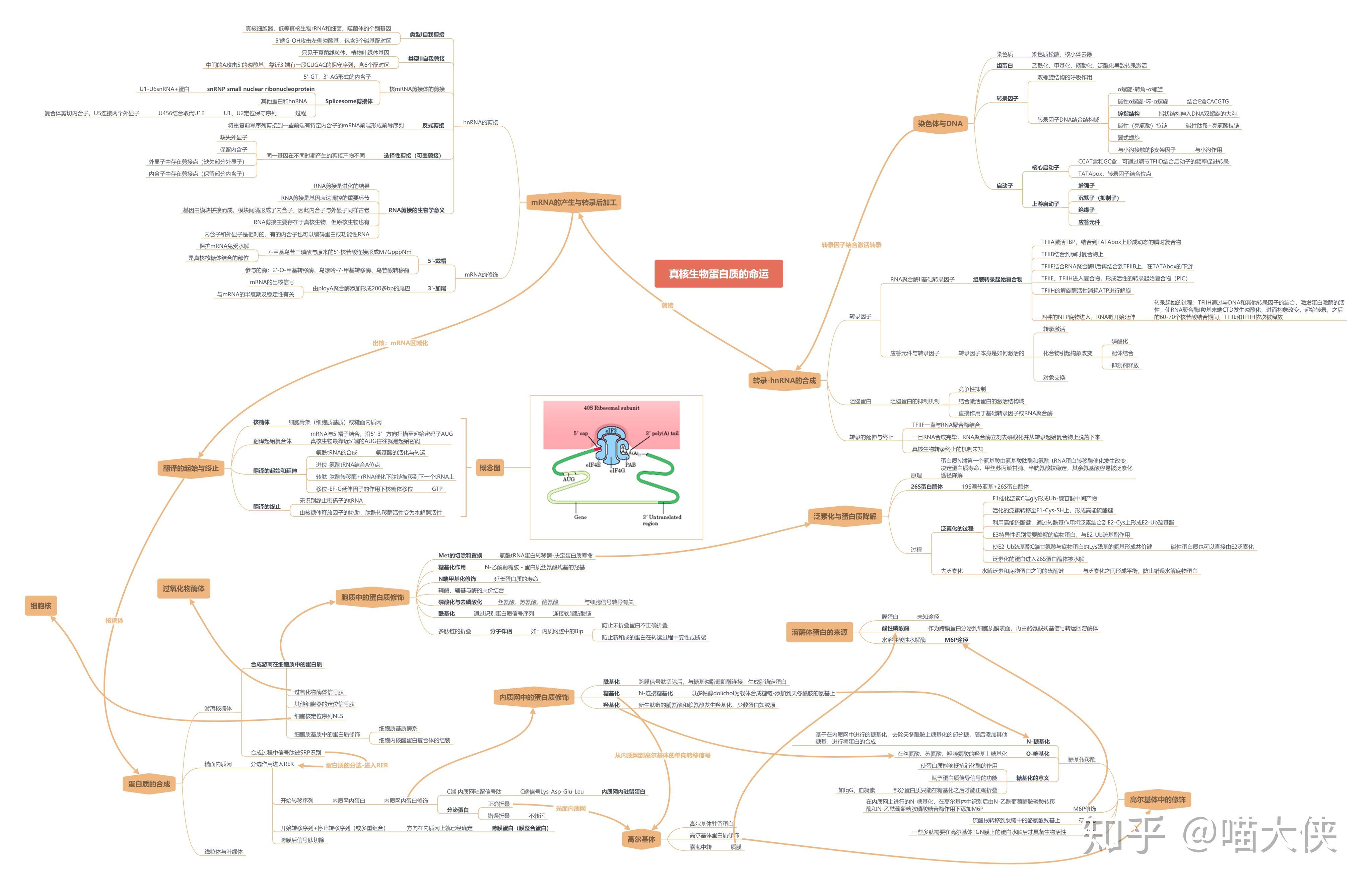
第三章 生物信息的传递(上)PPT已看
RNA的结构
- 由核酸、嘧啶组成
- 链自身折叠形成局部双螺旋
- 折叠形成三级
RNA的分类
RNA的功能
- 作为细胞内蛋白质合成的主要参与者
- 部分RNA可以作为核酶催化重要反应
- 参与基因表达的调控,与生物生长发育密切相关
- 在某些生物中,RNA是遗传物质
转录
RNA聚合酶与启动子DNA双链相互作用并与之相结合的过程
真核细胞需要转录调控因子才能与启动子结合
不需要引物,由RNA聚合酶催化,以NTP为底物连续合成RNA
在DNA上开始转录的第一个碱基为+1
形成9个核苷酸
通过启动子的时间代表启动子强弱
RNA聚合酶释放σ因子离开启动子,核心酶沿模板DNA链移动
内在终止子/不依赖ρ因子
终止位点上游一般存在一段富含GC碱基的二重对称区,其转录生成的mRNA容易互补形成发卡式结构
终止位点上游一般有4~8个A组成的序列,转录生成的mRNA 3&#39;末端中相应的有一连串U序列
依赖ρ因子
抗终止
依赖DNA的RNA聚合酶
特点
- 以DNA为模板
- 5&#39;→3&#39;连续合成
- 需要$$Mg^{2+}$$或$$Mn^{2+}$$
- 缺乏外切酶活性,无校正功能
- 不需要引物
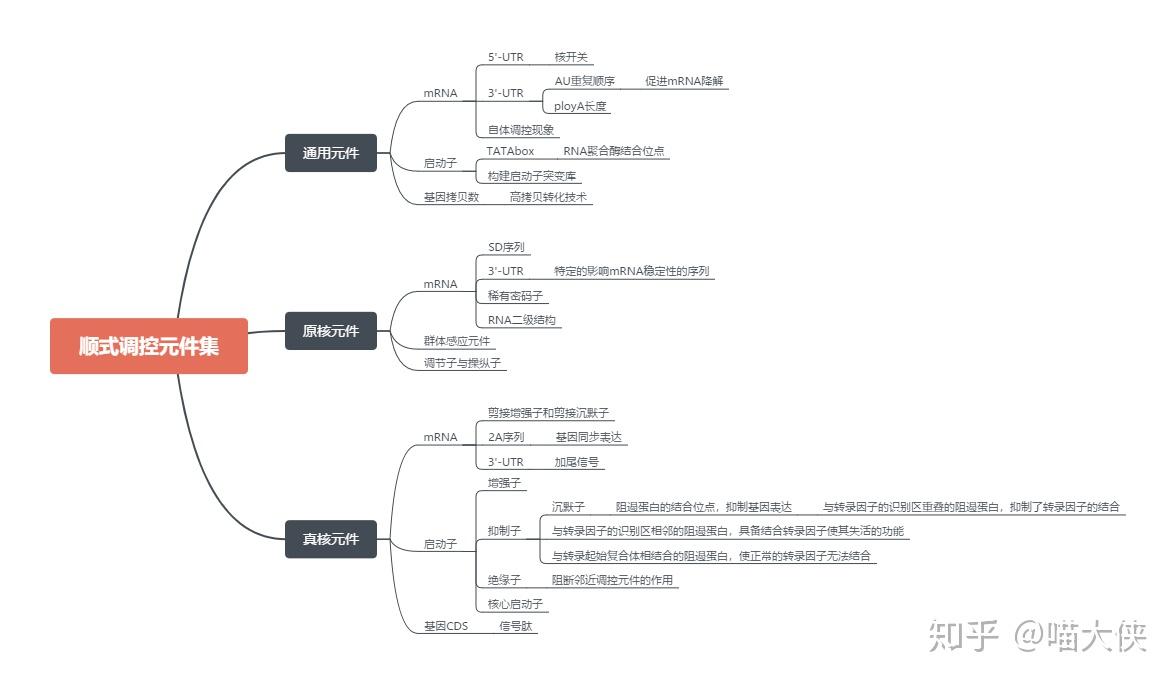
顺式作用元件--包括启动子,启动子上游元件,近段调控元件和增强子等远隔序列。
反式作用因子--能直接、间接辨认和结合转录上游区段DNA的蛋白质
基本转录因子--直接或间接结合RNA聚合酶的反式作用因子
起始点上游多数有共同的TATA序列,称为TATA盒。通常认为这就是启动子的核心序列启动子的功能既受DNA序列的影响,又受其构象的影响,推测RNA聚合酶并不直接识别碱基本身,而是通过氢键互补的方式加以识别(类似于酶与底物的结合)
启动子
- 启动子:一段位于结构基因 5&#39;端上游区的DNA序列。能活化RNA聚合酶,使之与模板DNA准确结合。
- 转录起始位点:与新生RNA链第一个核苷酸相对应DNA链上的碱基。通常为一个嘌呤。
- 转录单元:一段从启动子开始到终止子结束的DNA序列
-35序列(-35区)
- 保守性 $$T_{82}T_{84}G_{78}A_{65}C_{54}A_{45}$$
- -35序列的功能是σ因子初始识别并结合的位点,与RNA聚合酶全酶有很高的亲和性
- -35序列在很大程度上决定了启动子的强度
-10序列(Pribnow 区)
- TATAAT或稍有变化的形式,大致出现在-4至-13bp之间,保守性$$T_{80}A_{95}T_{45}A_{60}A_{50}T_{95}$$
- 决定转录方向,RNA聚合酶在此部位与DNA结合成稳定的复合物。
- Pribnow框中DNA序列在转录方向上解开双螺旋结构,形成开放型起始结构。
- RNA聚合酶牢固结合的位点,是启动子关键部位
最佳距离
- 原核生物中,-10区与-35区之间的距离大约是16-19bp,小于15或大于20都会降低启动子活性。
- 间隔的序列不严格,但是距离的大小可能是决定启动子强度的因素之一。
- 保持启动子这两段序列以及它们之间的距离十分重要,否则就会改变它所控制基因的表达水平
影响
- TATA区和上游启动子元件(CAAT区或GC区)的作用不同:
TATA区负责转录精确起始定位; CAAT区和GC区控制转录起始频率, CAAT区对转录起始频率的影响最大。
- TATA区和相邻的UPE区之间插入核苷酸会使转录减弱。
- 并不是每个基因的启动子区都包含这3种序列(TATA区、CAAT区、 GC区)
将TATA区上游的保守序列称为上游启动子(UPE)或上游激活序列(UAS)
增强子
- 远距离效应
- 无方向性
- 顺式调节
- 无物种和基因的特异性
- 具有组织特异性
- 有相位性
- 有的增强子可对外部信号产生反应
转录的抑制
- DNA模板功能抑制剂
- RNA聚合酶抑制剂
- 嘌呤和嘧啶类似物,抑制核酸前体的合成
真核/原核生物转录产物比较
原核生物RNA聚合酶
无法复制加载中的内容
转录起始时,原核生物RNA聚合酶与启动子的相互作用主要包括启动子区的识别、酶与启动子的结合及σ因子的结合与解离
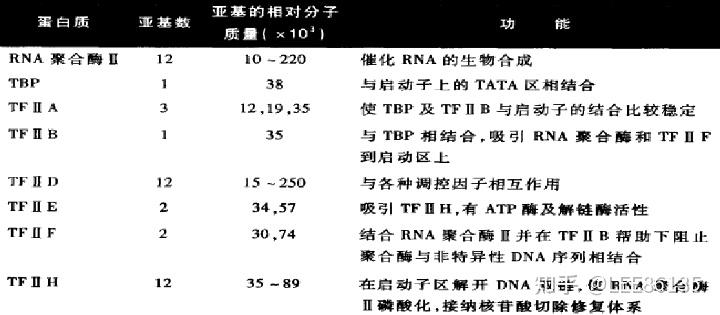
真核生物RNA聚合酶
真核生物RNA pol Ⅱ不与DNA分子直接结合,而需依靠众多的转录因子,形成转录起始前复合物(pre-initiationcomplex,PIC)
- 由TFIID中的TBP识别TATA盒,并在TAFs的协助下结合到启动子区,然后TFIIB与TBP结合,同时TFIIB也能与DNA结合,TFIIA可以稳定与DNA结合的TFIIB-TBP复合体;
- TFIIB-TBP复合体与RNA pol II-TFIIF复合体结合,此举可降低RNA pol II 与DNA的非特异部位的结合,协助RNA pol II 靶向结合启动子;
- TFIIE和TFIIH加入,形成闭合复合体,装配完成。
为了保证转录的准确性,不同基因需不同转录因子。
拼板理论(piecing theory) :少数几个反式作用因子(主要是可诱导因子和上游因子)之间互相作用,再与基本转录因子、RNA聚合酶搭配而有针对性地结合、转录相应的基因。可诱导因子和上游因子常常通过辅激活因子或中介子与基本转录因子、RNA聚合酶结合,但有时也可直接与基本转录因子、RNA聚合酶结合
RNA-pol前移处处都遇上核小体。转录延长过程中可以观察到核小体移位和解聚现象
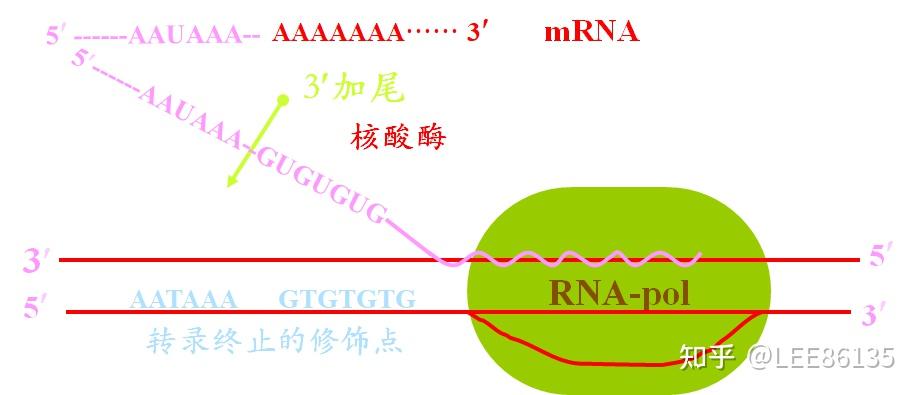
转录不是在poly A的位置上终止,而是超出数百个乃至上千个核苷酸后才停顿。
转录终止的修饰点--在读码框架的下游,常有一组共同序列AATAAA,再下游还有相当多的GT序列。
原核生物mRNA
- 许多原核生物mRNA可以多顺反子的形式存在
- 起始密码子通常是AUG(有时GUG、 UUG)
- 不需要转录后加工
- 原核生物mRNA的 5&#39;端无帽子结构,3&#39;端没有或只有较短的polyA结构
- 原核生物mRNA的半衰期短
真核生物mRNA
线粒体和叶绿体以AUG、AUU、AUA 为起始密码子
- 需要转录后加工
- mRNA的5&#39;端存在“帽子”结构
- 绝大多数真核生物mRNA具有多聚A尾巴
- 加工产物
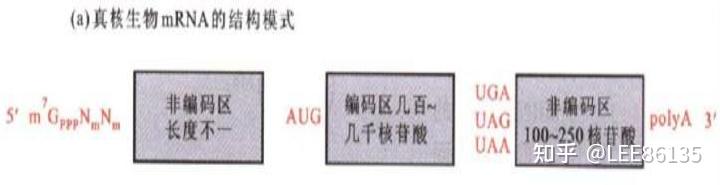
真核生物mRNA主要的修饰方式--剪接、剪切、修饰、添加
5&#39;端和3&#39;端(首、尾部)的修饰
大多数真核mRNA的5&#39;末端有7-甲基鸟嘌呤的帽结构。由加帽酶和甲基转移酶催化完成
帽子结构
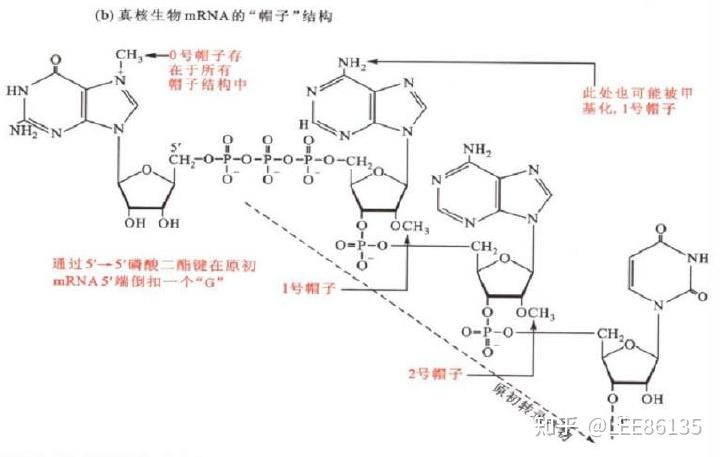
第一个G的N7位甲基化称为帽0--5&#39;m7GpppNp-3&#39;
核糖或第二碱基甲基化称为帽1--5&#39;m7GpppNmp-3&#39; (R1=CH3)/5&#39; Gpppm7Nmp-3&#39; (第二碱基的N7位)
2个核糖的R1、R2甲基化称为帽2--m7GpppN1mpN2mp- ($$R1=CH_3、R2=CH_3$$)
多个位点甲基化称为帽3
帽子结构的作用
- 帽子结构可使mRNA免遭核酸酶的破坏。
- 有帽子结构的mRNA更容易被蛋白质合成的起始因子所识别,从而促进蛋白质的合成。
polyA尾巴
尾部修饰和转录终止同时进行
polyA是mRNA由细胞核进入细胞质所必需的形式,poly A的有无与长短,是维持mRNA作为翻译模板的活性,以及增加mRNA本身稳定性的因素。
一般真核生物在胞浆内出现的mRNA,其poly A长度为100至200个核苷酸之间,也有少数例外。
前体mRNA分子的断裂和加多聚腺苷酸尾是多步骤过程--在转录后,由内切酶切开mRNA3&#39;端特定部位,然后由poly A 聚合酶催化多聚腺苷酸反应
mRNA刚进入进入胞质时,poly A尾巴较长,随时间推移将变短直至消失,随后mRNA将降解
对hnRNA进行剪接
mRNA的剪接
共有序列的识别和复合体的组装;进行断裂和连接反应进而改变RNA底物的结构
通过折叠形成鸟苷G结合口袋
腺苷酸A形成套索状产物
hnRNA--mRNA前体
成熟的mRNA,被转运到核糖体,指导蛋白质翻译
GU-AG法则--内含子的两端边界存在共同的序列。此规则不适合于线粒体和叶绿体基因的内含子,也不适合于tRNA和rRNA的内含子
根据基因的类型和剪接的方式,通常把内含子分为4类:
I主要存在于线粒体,叶绿体及某些低等真核生物的rRNA;
II也发现于线粒体,叶绿体,转录产物是mRNA;
III是常见的形成套索结构后剪接,大多数mRNA基因有此类内含子;
IV是tRNA基因及其初级转录产物中的内含子,剪接过程需酶及ATP
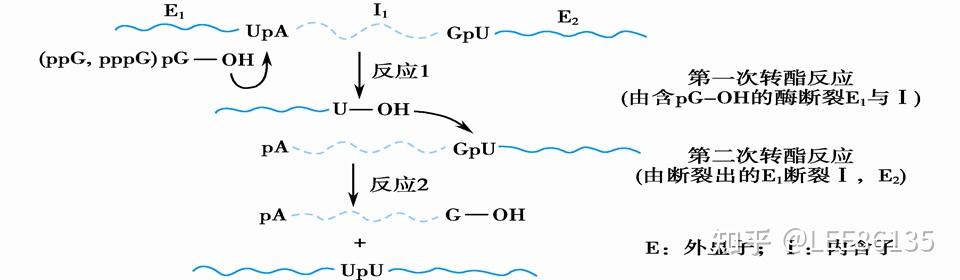
第一次:内含子和外显子交界处;第一次反应是由剪接体的辅助因子鸟嘌呤的羟基对UpA的磷酸酯健的酯健进行第一次转酯反应。转酯反应的结果把外显子1切掉。
第二次:外显子1,内含子,和外显子2交界处;外显子1切掉以后3&#39;端有羟基,3&#39;端的羟基作为催化集团对GpU的磷酸酯健的酯健再来一次转酯反应,剩下的内含子切掉的。
- 转酯反应是由一个称为剪接体的大复合体介导的。剪接体中包含约150种蛋白和5种小RNA,大小类似核糖体。
- RNA和蛋白质都参与共有序列的识别。剪接体的多数功能是由RNA分子执行的。
- RNA分子识别内含子与外显子交界序列,亲自催化剪接。
- 5种核内小RNA(snRNA) :U1,U2,U4,U5,U6 (序列中富含U)。
- (小的核内核糖核蛋白)snRNP,RNA与蛋白质形成RNA-蛋白质复合物
- AG前一位核苷酸可以影响剪切效率:CAG=UAG>AAG>GAG
snRNP在剪接中的功能
- 识别5&#39;剪接位点和分支点;
- 把这2个位点集结到一起;
- 催化或协助催化剪接和连接反应
内含子在体外无需蛋白质协助可自身剪接,在体内需要蛋白质组分激活
真核生物tRNA加工
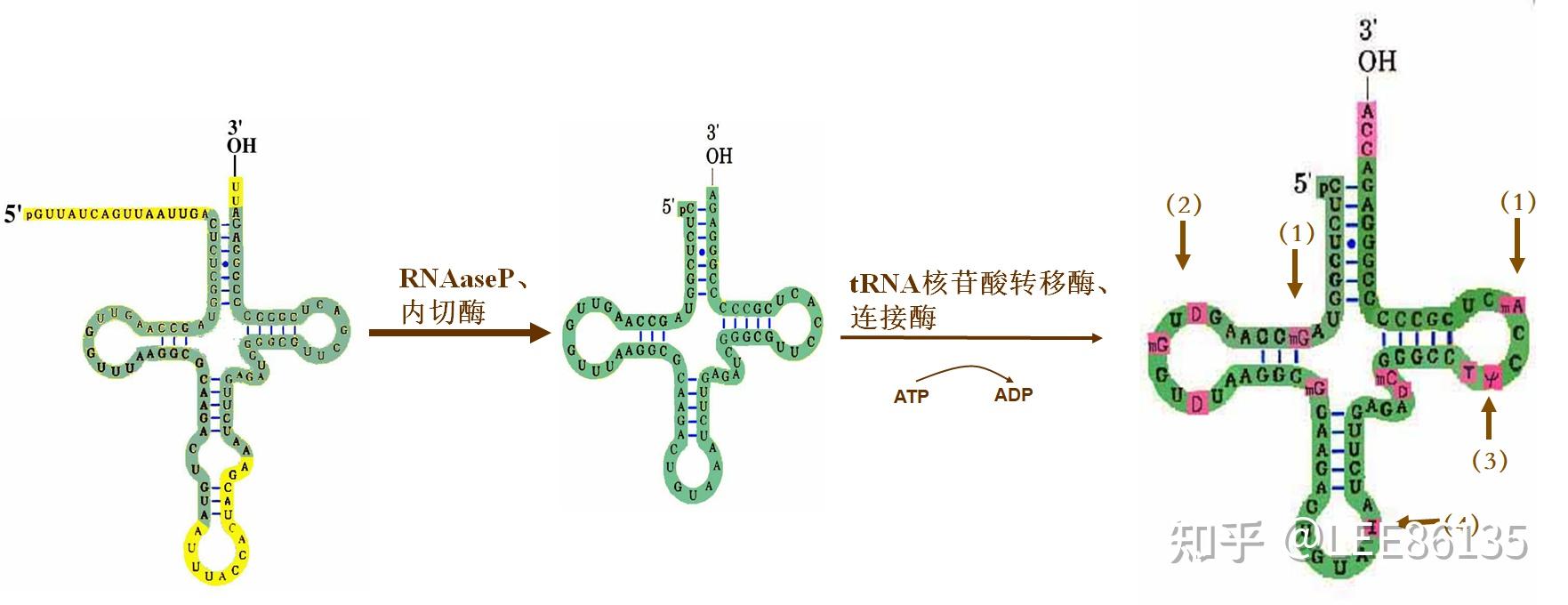
内含子切除不能够自行催化,必须有内切酶和核酸酶p来把它切掉
- 甲基化--如A→$$A^m$$
- 还原反应--如U→DHU
- 核苷内的转位反应--如U→ψ
- 脱氨反应--如A→I
- 转录与翻译的时空差
原核生物转录和翻译发生在同一个细胞空间,几乎同步
RNA的编辑
分化加工--基因的编码序列经过转录加工后多用途化
位点特异性脱氨基作用
引导RNA指导的尿嘧啶插入或删除
意义
RNA再编码
- 核糖体程序性+1/-1移位(Shift)
- 核糖体跳跃
- 终止子通读(UGA:硒代半胱氨酸、UAG:吡咯赖氨酸)
RNA的化学修饰
- 甲基化
- 去氨基化
- 硫代
- 碱基的同分异构化
- 二价键的饱和化
- 核苷酸的替代
核酶
一类具有催化功能的RNA分子,通过催化靶位点RNA链中的磷酸二酯键的断裂,特异性地剪切底物RNA分子,从而阻断基因的表达
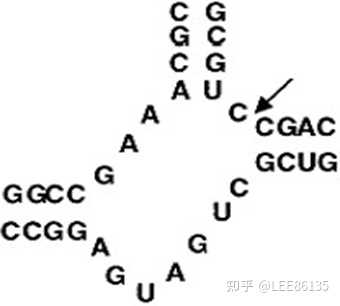
通常为60个核苷酸左右,同一个分子上包括有催化部分和底物部分,催化部分和底物部分组成锤头状结构
45S的转录产物经过剪切后生产核糖体小亚基的18SrRNA和5.8SrRNA、28SrRNA
作用方式
- 剪切型--切除RNA前体多余部分
- 剪接型--切除RNA前体的内含子部分,把不连续的外显子连接起来
意义
- 突破了酶的概念,是一种自体催化。
- 揭示了内含子自我剪接的奥秘,促进了RNA研究。
- 为生命起源和分子进化提供了新依据
RNA在生物进化中的地位
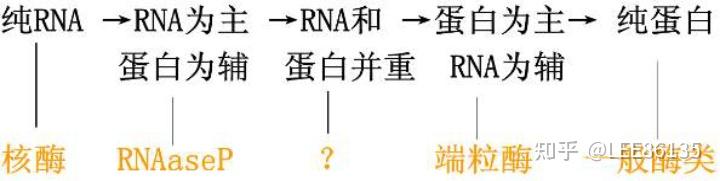
酶组成的进化 生物体组成的进化
第四章 生物信息的传递(下)PPT已看
确认三联子方法和现象
- 移码突变
- 烟草坏死病外壳蛋白亚基400aa/1200bp
- 核苷酸的插入或删除实验
Crick提出遗传密码的特点
- 3个碱基编码一个氨基酸
- 密码不重叠
- 氨基序列从固定起点解读
- 密码是简并的
破译密码子的方法
- 体外翻译系统的建立
- 核酸的人工合成技术的发展
- 核糖体结合技术
- 以均聚物、随机共聚物和特定序列的共聚物为模板指导多肽的合成
遗传密码的性质
- 连续性
- 简并性
- 普遍性与特殊性
- 密码子与反密码子相互作用
摆动假说
解释了反密码子中某些稀有成分的配对与许多氨基酸有2个以上密码子的现象
tRNA
一个tRNA能识别的密码子数量由反密码子的第一位碱基性质决定

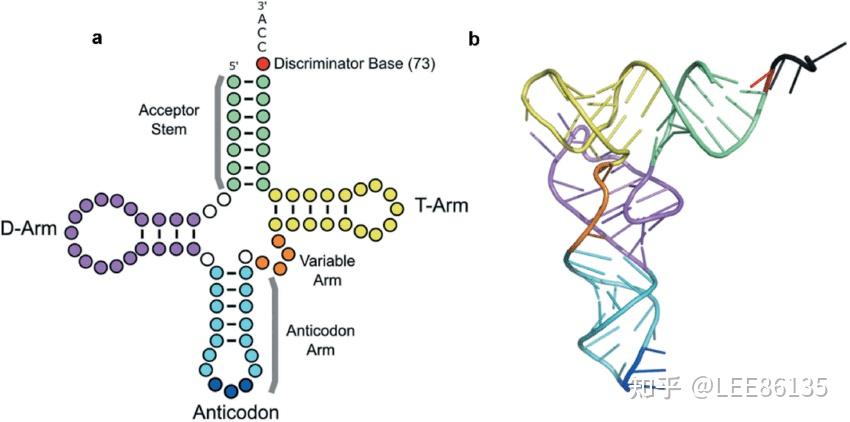
- 受体臂:3&#39;端最后三个碱基为CCA
- TψC臂:ψ表示拟尿嘧啶,为tRNA分子所拥有的不常见核苷酸
- 反密码子臂:
- D臂:含有二氢尿嘧啶
特点
- 稀有碱基很丰富,约70余种
- 所有tRNA都能与核糖体A位点(氨酰-tRNA结合位点)、P位点(肽酰-tRNA结合位点)结合
- tRNA能被翻译辅助因子识别
二级结构中未配对碱基间形成氢键引发三级结构的形成
种类
起始tRNA和延伸tRNA
同工tRNA几个代表相同氨基酸的tRNA
校正tRNA(改变反密码子区)--无义突变的校正tRNA,错义突变的校正tRNA
无义突变--原密码子变为终止子,合成无功能或无意义的多肽
错义突变--变为另一氨基酸的密码子
校正tRNA校正时要与正常tRNA竞争密码子
校正基因的效率由反密码子与密码子的亲和力和细胞中浓度及其他参数决定
氨酰-tRNA合成酶
核糖体
小亚基大亚基
核糖体蛋白
核糖体RNA
三个tRNA结合位点E-P-A
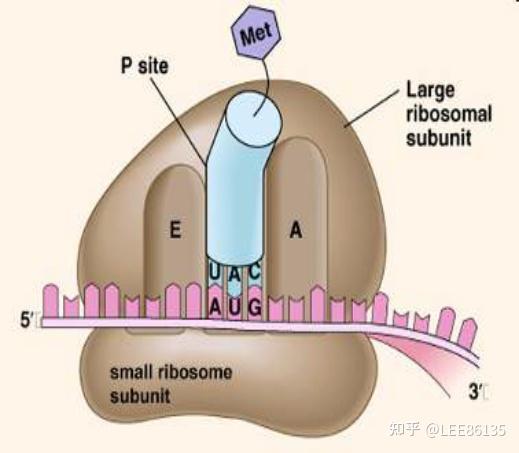
真核生物RNA3/5,蛋白质2/5
原核生物RNA2/3,蛋白质1/3
功能
- 核糖体包括5个以上活性中心
- 核糖体小亚基负责对模板mRNA进行序列特异性识别,可与GTP、起始tRNA结合
- 大亚基负责携带AA-tRNA、肽键的形成、AA-tRNA与肽链的结合等,具有转肽酶活性,将P位肽酰基转移给A位上的氨基酰tRNA;具有起始因子、延长因子、及释放因子的结合部位;具有GTPase活性,能够水解GTP获得能量
- 核糖体在体内及体外都可解离为亚基或结合成70S/80S颗粒
蛋白质合成的生物学机制
mRNA的结构
从mRNA 5&#39;-端起始密码子AUG到3&#39;-端终止密码子之间的核苷酸序列,称为开放阅读框架(open reading frame, ORF)
开放阅读框通常代表某个基因的编码序列
遗传学将编码一个多肽的遗传单位称为顺反子(cistron)。
原核细胞中数个结构基因常串联为一个转录单位,转录生成的mRNA可编码几种功能相关的蛋白质,为多顺反子(polycistron) 。
真核mRNA只编码一种蛋白质,为单顺反子(single cistron)
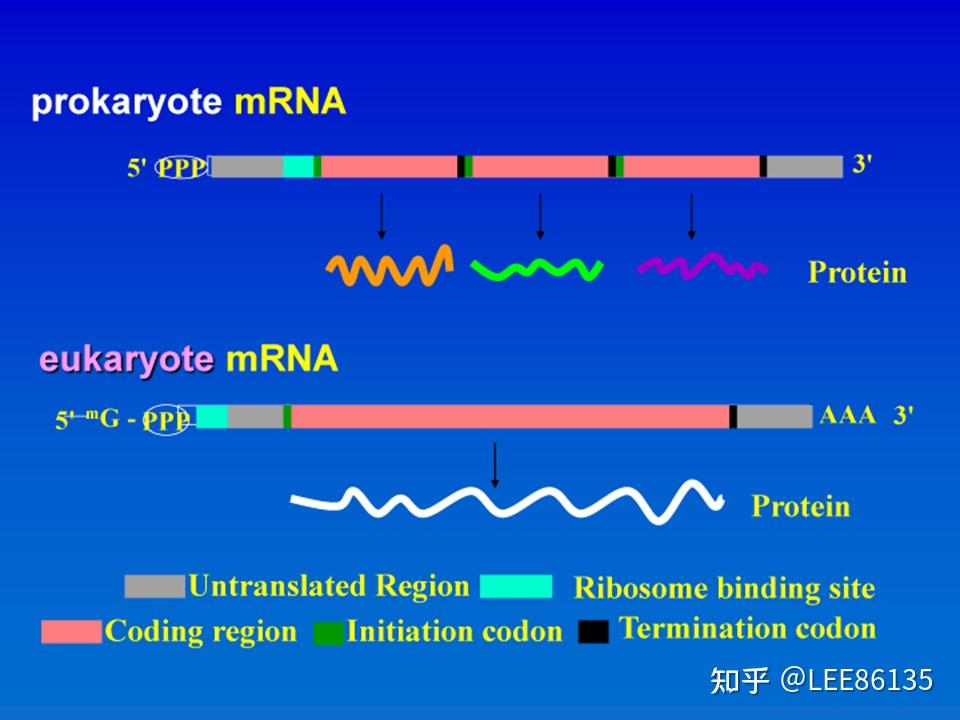
起始密码(initiation codon):AUG
终止密码(termination codon):UAA,UAG,UGA
无法复制加载中的内容
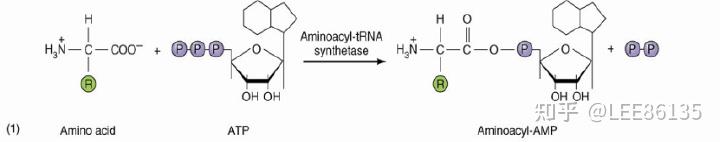
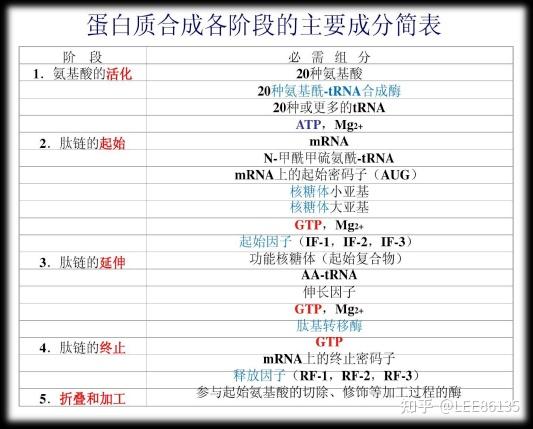
氨基酸活化
酶
氨基酰-tRNA合成酶(aminoacyl-tRNA synthetase),催化氨基酸的活化;
转肽酶(peptidase),催化核糖体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离;
转位酶(translocase),催化核糖体向mRNA3&#39;-端移动一个密码子的距离,使下一个密码子定位于A位
蛋白质因子
起始因子
促进核糖体小亚基与起始tRNA及模板mRNA结合
原核生物中存在3种起始因子,分别称为IF1-3。真核生物中存在9种以上起始因子(eIF)
延长因子(EF)
促使氨基酰tRNA进入核糖体的受体,并可促进移位过程
原核生物中存在3种延长因子(EF-Tu,EF-Ts,EF-G),真核生物中存在2种(EF1,EF2)。
核糖体mRNA移动和肽酰t-RNA的移位是偶联的
EF-Tu,EF-Ts促进AA-tRNA进入A位,EF-G促进移位和卸载tRNA的释放
释放因子(RF)
识别终止密码,协助多肽链的释放
原核生物中有4种,在真核生物中只有1种。
供能物质和无机离子
多肽链合成时,需ATP、GTP作为供能物质,并需$$Mg^{2+}、K^+$$参与。
氨基酸活化时需消耗2分子高能磷酸键,肽键形成时又消耗2分子高能磷酸键,故缩合一分子氨基酸残基需消耗4分子高能磷酸键
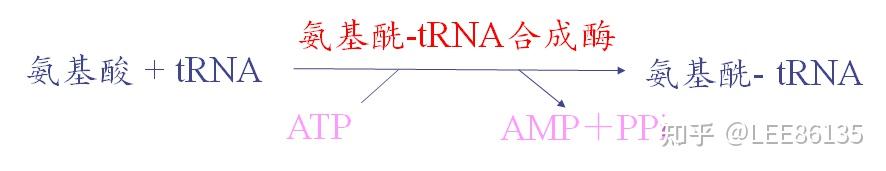
氨基酰-tRNA合成酶的特点
氨基酰-tRNA合成酶对底物氨基酸和tRNA都有高度特异性。
氨基酰-tRNA合成酶具有校正活性:能将错误的结合的氨基酸水解释放,再换上与密码子相对应的氨基酸
氨基酰-tRNA的表示方法:$$Ala-tRNA^{Ala}$$
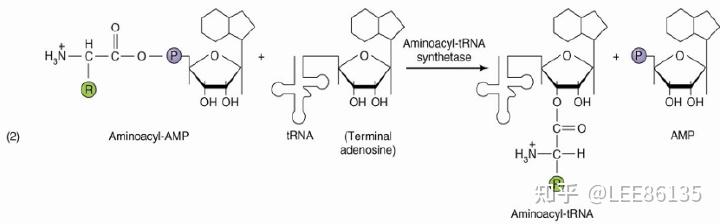
能够识别mRNA中5′端起始密码AUG的tRNA是一种特殊的tRNA,称为起始tRNA。
在原核生物和真核生物中,均存在另一种携带甲酰甲硫氨酸或甲硫氨酸的tRNA,识别非起动部位的甲酰甲硫氨酸或甲硫氨酸密码,AUG。
肽链的起始
起始因子是在蛋白质合成的起始阶段与核糖体小亚基结合,帮助形成起始复合物的蛋白因子。这些蛋白因子只与核糖体小亚基结合,当小亚基与大亚基结合成核糖体时,起始因子即被释放。
原核生物翻译的起始
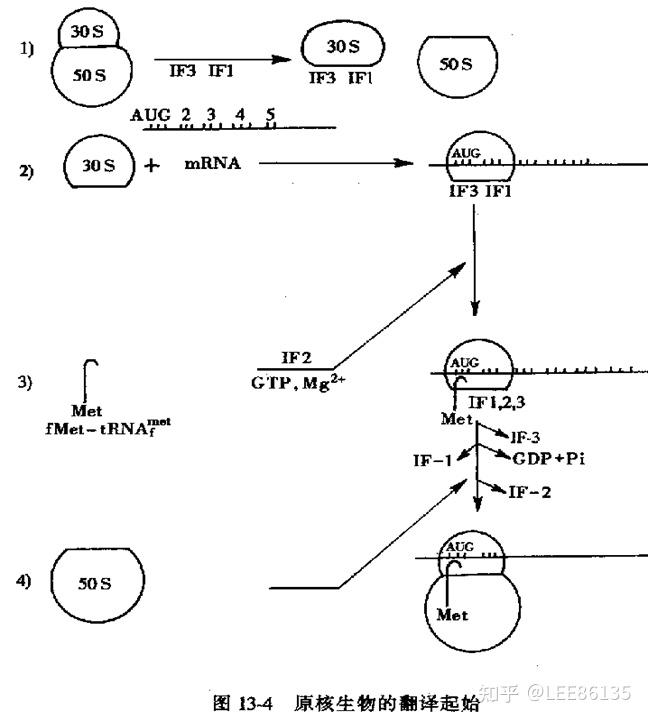
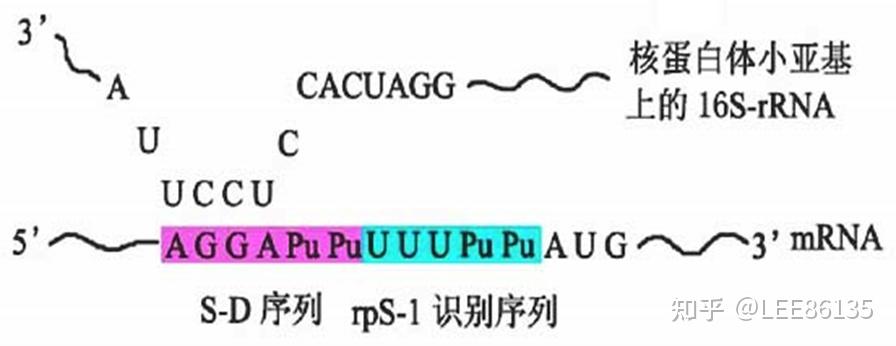
- 30S小亚基与翻译起始因子IF-1,IF-3结合,通过SD序列与模板结合。在各种mRNA起始AUG上游约8~13核苷酸部位,存在一段由4~9个核苷酸组成的一致序列,富含嘌呤碱基,如-AGGAGG-,称为Shine-Dalgarno序列(S-D序列),又称核蛋白体结合位点(ribosomal binding site, RBS)。
- 在IF-2和GTP帮助下,fMet进入小亚基的P位,tRNA上的反密码子与mRNA上的起始密码子配对
- 带有tRNA、mRNA、三个翻译起始因子的小亚基复合物与50S大亚基结合,GTP水解并释放翻译起始因子
肽链的延伸
指根据mRNA下一组遗传密码指导,使相应氨基酰-tRNA进入核糖体A位。
此步骤需GTP,Mg2+,和EF参与
在转肽酶的催化下,将P位上的tRNA所携带的甲酰甲硫氨酰基或肽酰基转移到A位上的氨基酰-tRNA上,与其α-氨基缩合形成肽键。
此步骤需Mg2+,K+。
P位上已失去甲酰甲硫氨酰基或肽酰基的tRNA从核糖体上脱落
由转位酶(translocase)催化,其活性存在于EF-G
此步骤需GTP和Mg2+参与。
核糖体向mRNA的3&#39;- 端滑动相当于一个密码的距离,同时使肽酰tRNA从A位移到P位,卸载的tRNA进入E位。
核糖体的A位留空,与下一个密码相对应的氨基酰-tRNA即可再进入,重复以上循环过程,使多肽链不断延长
肽链的终止
- 识别:RF识别终止密码,进入核糖体的受位。
- 水解:RF使转肽酶变为水解酶,多肽链与tRNA之间的酯键被水解,多肽链释放。
- 解离:通过水解GTP,使核糖体与mRNA分离,tRNA、RF脱落,核糖体解离为大、小亚基
蛋白质前体的加工
无法复制加载中的内容
- 二硫键的形成。
- 特定氨基酸的修饰。氨基酸侧链的修饰包括:
• 磷酸化(如核糖体蛋白质)
• 糖基化(如各种糖蛋白)
• 甲基化(如组蛋白、肌肉蛋白质)
• 乙基化(如组蛋白)
• 羟基化(如胶原蛋白)和羧基化等
- 切除新生链中非功能片段
- 空间结构修饰--亚基聚合、辅基连接、疏水脂键的共价连接
蛋白质折叠
- 新生多肽链必须经过正确的折叠,才能形成动力学和热力学稳定的三维构象,从而表现出生物学活性或功能
- 多肽链的折叠是一个复杂的过程,新生肽链一般首先折叠成二级结构,再进一步折叠盘绕成三级结构
- 分子伴侣是能在细胞内辅助新生肽链正确折叠的一类蛋白质,主要有热休克蛋白、伴侣素、触发因子等
热休克蛋白
主要包括HSP70、HSP40、GrpE三族

大肠杆菌的HSP40 (Dna J)可激活Dna K中的ATP酶,生成稳定的Dna J-Dna K-ADP-被折叠蛋白质复合物,以利于Dna K发挥分子伴侣作用。在ATP存在的情况下,Dna J和Dna K的相互作用能抑制蛋白质的聚集。
Grp E,核苷酸交换因子,与Dna K的ATP酶结构域结合,使Dna K的构象发生改变、ADP从复合物中释放出来并由ATP代替ADP,从而控制Dna K的ATP酶活性。
热休克蛋白促进蛋白质折叠的基本作用--结合保护待折叠多肽片段,再释放该片段进行折叠。形成HSP70和多肽片段依次结合、解离的循环。
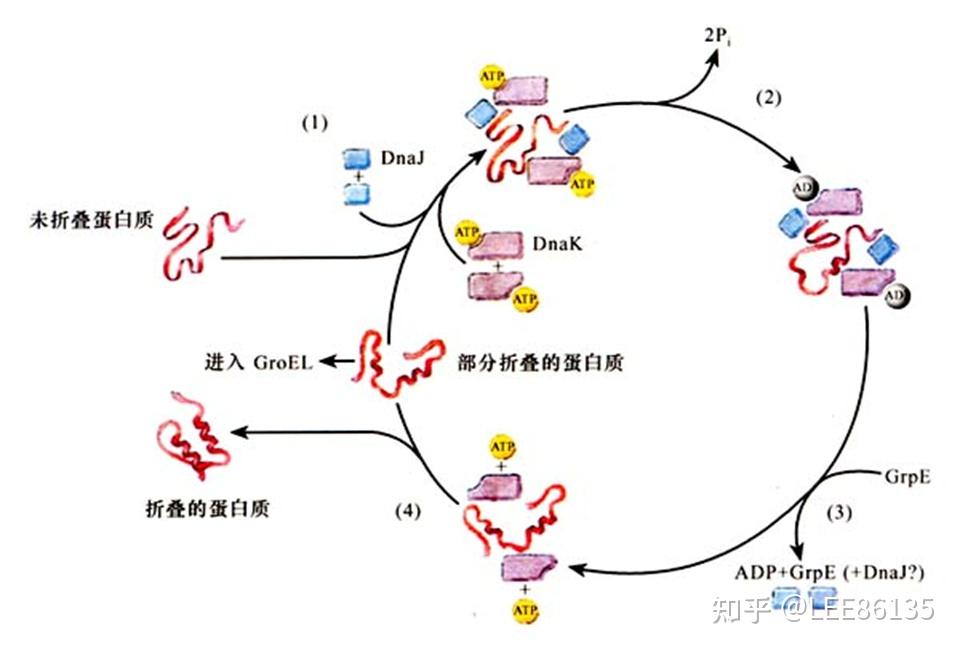
- HSP40结合待折叠多肽片断
- 将多肽导向HSP70-ATP复合物
- 产生稳定的HSP40-HSP70-ADP-多肽复合物
- 核苷酸交换因子GrpE与HSP70作用,促进ATP交换ADP
- 复合物解离,释出多肽链片断
- 多肽链片断正确折叠
人类细胞中HSP蛋白质家族可存在于胞浆、内质网腔、线粒体、胞核等部位,涉及多种细胞保护功能:如使线粒体和内质网蛋白质保持未折叠状态而转运、跨膜,再折叠为功能构象;通过类似上述机制,避免或消除蛋白质变性后因疏水基团暴露而发生的不可逆聚集,以利于清除变性或错误折叠的多肽中间物等。
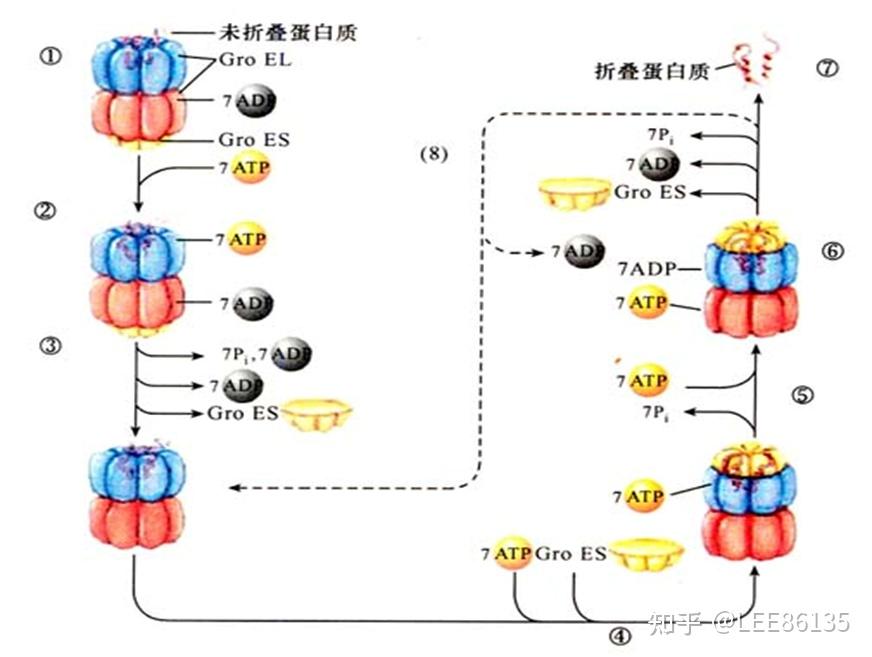
伴侣蛋白
主要作用是为非自发性折叠蛋白质提供能折叠形成天然空间构象的微环境
Gro EL-Gro ES反应循环
多肽链内或肽链之间二硫键的正确形成对稳定分泌蛋白、膜蛋白等的天然构象十分重要,这一过程主要在细胞内质网进行
蛋白二硫键异构酶在内质网腔活性很高,可在较大区段肽链中催化错配二硫键断裂并形成正确二硫键连接,最终使蛋白质形成热力学最稳定的天然构象
肽链中肽酰-脯氨酸间形成的肽键有顺反两种异构体,空间构象明显差别。肽酰-脯氨酰顺反异构酶可促进上述顺反两种异构体之间的转换。
肽酰-脯氨酰顺反异构酶是蛋白质三维构象形成的限速酶,在肽链合成需形成顺式构型时,可使多肽在各脯氨酸弯折处形成准确折叠。
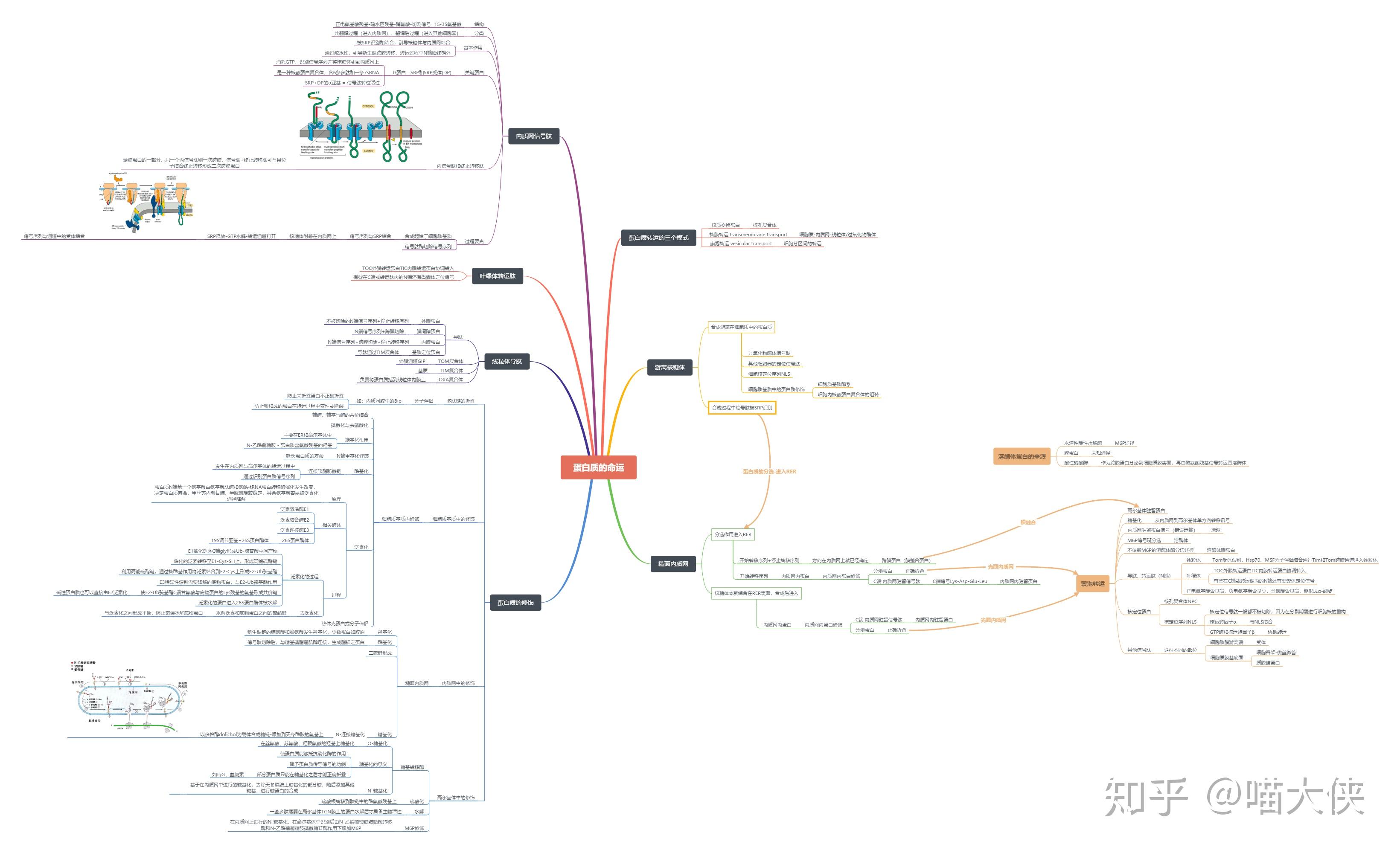
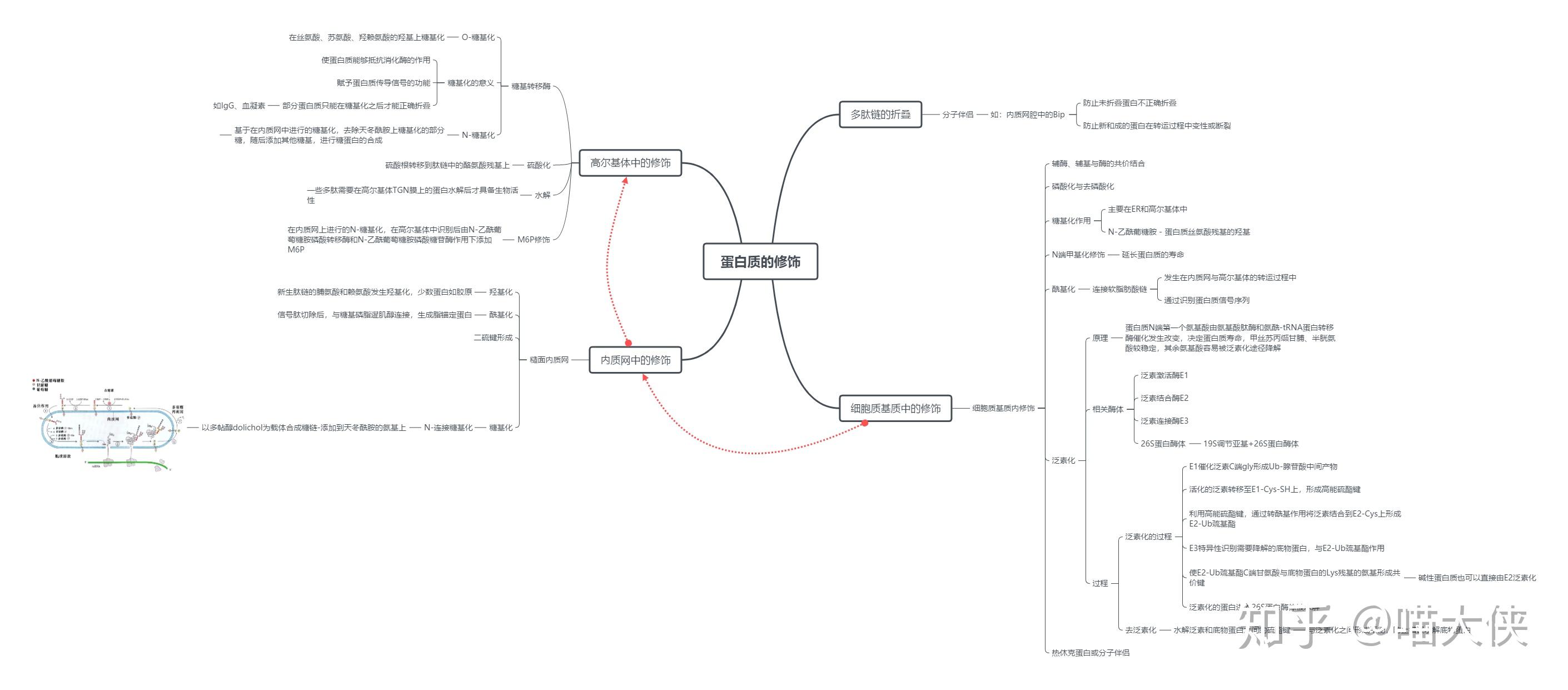
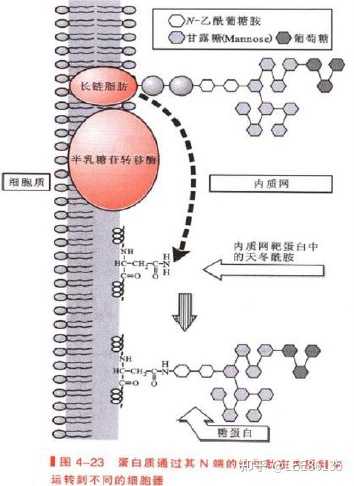
蛋白质合成后的靶向输送
蛋白质的靶向输送与翻译后修饰过程同步进行
信号序列--所有靶向输送的蛋白质结构中存在分选信号,主要为N末端特异氨基酸序列,可引导蛋白质转移到细胞的适当靶部位
信号肽--各种新生分泌蛋白的N端有保守的氨基酸序列
信号肽约13~36氨基酸残基,分碱性区、疏水核心区、加工区三个区段。
信号肽的N端为带正电荷的氨基酸残基(赖、精);中间为疏水的核心区(亮、异亮等),而C端由小分子氨基酸残基(丝、甘、丙等)组成,可被信号肽酶识别并裂解。
蛋白质合成的抑制剂
抗生素
- 阻止mRNA与核糖体结合,抑制转肽酶、阻断肽链延长(氯霉素)
- 阻止AA-tRNA与核糖体结合,抑制氨基酰-tRNA与小亚基结合 (四环素类)
- 干扰AA-tRNA与核糖体结合而产生错读,改变构象引起读码错误、抑制起始(链霉素能干扰fMet-tRNA与核糖体的结合、新霉素、卡那霉素等)
- 提前释放肽链来抑制蛋白质的合成(嘌呤霉素)
- 抑制转肽酶、阻断肽链延长(放线菌酮)
干扰素
干扰素(interferon, IFN)是真核细胞被病毒感染后分泌的一类具有抗病毒作用的蛋白质,可抑制病毒的繁殖。
干扰素分为α-(白细胞)型、β-(成纤维细胞)型和γ-(淋巴细胞)型三大类,每类各有亚型,分别具有其特异作用。
机制
- 在某些病毒双链RNA存在时,能诱导特异的蛋白激酶活化,该活化的蛋白激酶使eIF-2磷酸化而失活,从而抑制病毒蛋白质合成;
- 能与双链RNA共同活化特殊的2&#39;-5&#39;寡聚腺苷酸(2&#39;-5&#39;A)合成酶,催化ATP聚合,生成单核苷酸间以2&#39;-5&#39;磷酸二酯键连接的2&#39;-5&#39;A,经2&#39;-5&#39;A活化核酸内切酶RNase L,后者可降解病毒mRNA,从而阻断病毒蛋白质合成
毒素
白喉毒素--ADP糖基化共价修饰真核生物延长因子eEF-2,使其失活
植物毒素蓖麻蛋白--真核生物大亚基失活
蛋白质转运机制
翻译-转运同步机制
蛋白质的合成和运转同时发生。分泌蛋白质大多是以同步机制运输的。
- 蛋白质定位的信息存在于该蛋白质自身结构中,并且通过与膜上特殊受体的相互作用得以表达
- 蛋白质跨膜转运信号也是由mRNA编码的。
- 在起始密码子后,有一段编码疏水性氨基酸序列的RNA区域,被称为信号序列
- 10~15个疏水氨基酸
- N端有带正电荷的氨基酸。
- C端接近切割点处代数个极性氨基酸;离切割点最近的氨基酸往往带有很短的侧链(Ala, Gly)
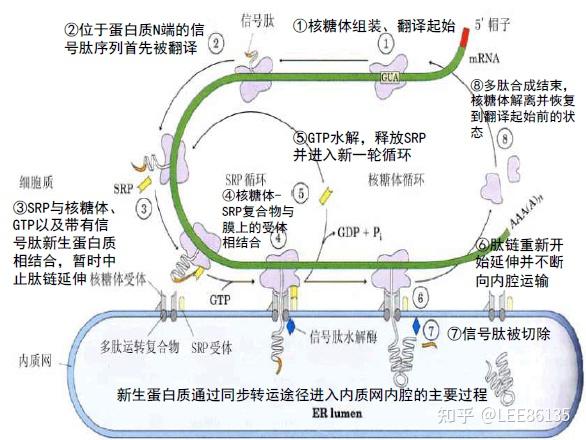
该机制说明
- 完整的信号肽是保证蛋白质转运的必要条件。
- 仅有信号肽还不足以保证蛋白质转运的发生,还要求转运蛋白质在信号序列以外的部分有相应的结构变化
- 信号序列的切除并不是转运所必需的。
- 并非所有的转运蛋白都有可降解的信号肽。
翻译后转运机制
蛋白质从核糖体上释放后才发生转运。在细胞器发育过程中,由细胞质进入细胞器的蛋白质大多是以翻译后转运机制运输的
线粒体蛋白质跨膜转运
特征
- 通过线粒体膜的蛋白质在转运前大多数以前体形式存在,它由成熟蛋白质和位于N端的一段前导肽共同组成,前导肽约含20~80个氨基酸残基
- 蛋白质通过线粒体膜运转是一种需要能量的过程。
- 蛋白质通过线粒体膜运转时,首先由外膜上的Tom受体复合蛋白识别与分子伴侣相结合的待运转多肽,通过Tom和Tim组成的膜通道进入线粒体内腔
前导肽(leader peptide)的作用与性质
拥有前导肽的线粒体蛋白质前体能够跨膜运转进入线粒体,在这一过程中前导肽被水解,前体转变为成熟蛋白,则失去继续跨膜能力。因此,前导肽对线粒体蛋白质的识别和跨膜运转起着关键作用
特性
- 带正电荷的碱性氨基酸(特别是精氨酸)含量较为丰富,它们分散于不带电荷的氨基酸序列之间
- 缺少带负电荷的酸性氨基酸
- 羟基氨基酸(特别是丝氨酸)含量较高
- 有形成两亲(既有亲水又有疏水部分)а-螺旋结构的能力
叶绿体蛋白质的跨膜转运
叶绿体多肽在胞质中的游离核糖体上合成后脱离核糖体并折叠成具有三级结构的蛋白质分子,多肽上某些特定位点结合于只有叶绿体膜上才有的特异受体位点。叶绿体定位信号肽一般有两个部分,第一部分决定该蛋白质能否进入叶绿体基质,第二部分决定该蛋白能否进入类囊体
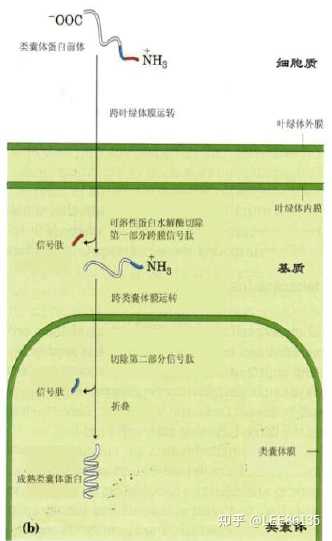
特点
- 活性蛋白水解酶位于叶绿体基质内。
- 叶绿体膜能够特异地与叶绿体蛋白的前体结合。
- 叶绿体前体内可降解序列因植物和蛋白质种类不同而表现出明显的差异
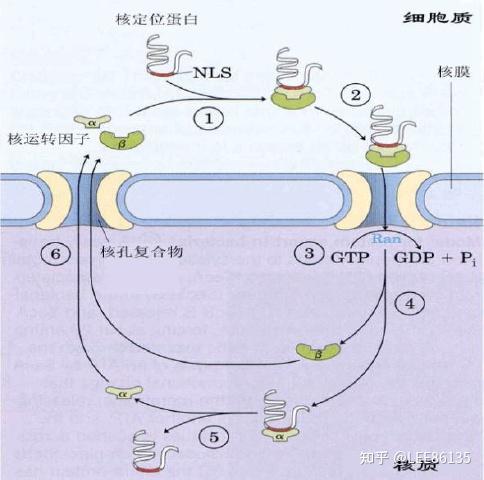
核定位蛋白的转运机制
为了核蛋白的重复定位,这些蛋白质中的信号肽——被称为核定位序列(NLS)一般都不被切除。NLS可以位于核蛋白的任何部位
蛋白质向核内运输过程需要核运转因子α、β和一个低分子量GTP酶(Ran)参与
蛋白质的修饰、降解
- 大肠杆菌蛋白的降解需要一个依赖于ATP的蛋白酶(Lon)。细胞中出现错误的蛋白质或半衰期很短的蛋白质时,该酶被激活。
- 真核生物蛋白降解依赖于泛蛋白(ubiquitin)
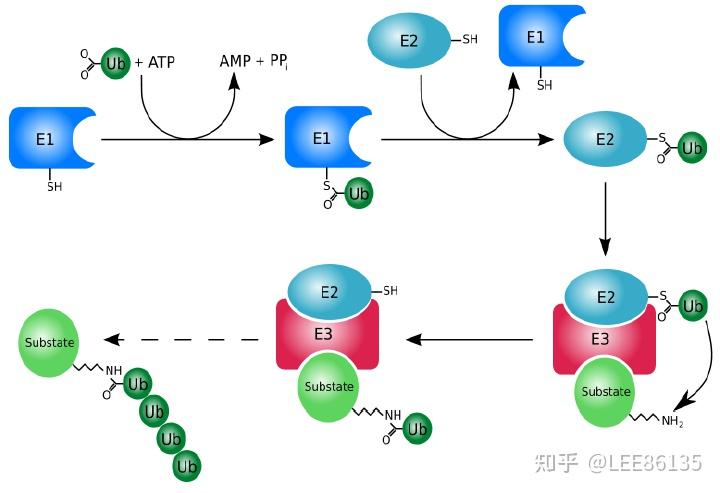
E1酶负责活化泛蛋白Ub
E2酶通过转硫醇作用从E1酶处获得泛蛋白,并将其与底物蛋白相结合
E3酶将泛蛋白与底物连接
泛蛋白降解意义
- 清除错误蛋白质,对细胞生长周期、DNA复制以及染色体结构都有重要的调控作用
- 对理解细胞的许多生理过程和新药的开发具有重要意义
蛋白质的稳定性研究
成熟蛋白N端的第一个氨基酸(除已被切除的N端甲硫氨酸之外,但包括翻译后修饰产物)在蛋白的降解中的影响很大。当某个蛋白质的N端是甲硫氨酸、甘氨酸、丙氨酸、丝氨酸、苏氨酸和缬氨酸时,表现稳定。其N端为赖氨酸、精氨酸时,表现最不稳定,2-3min就被降解了
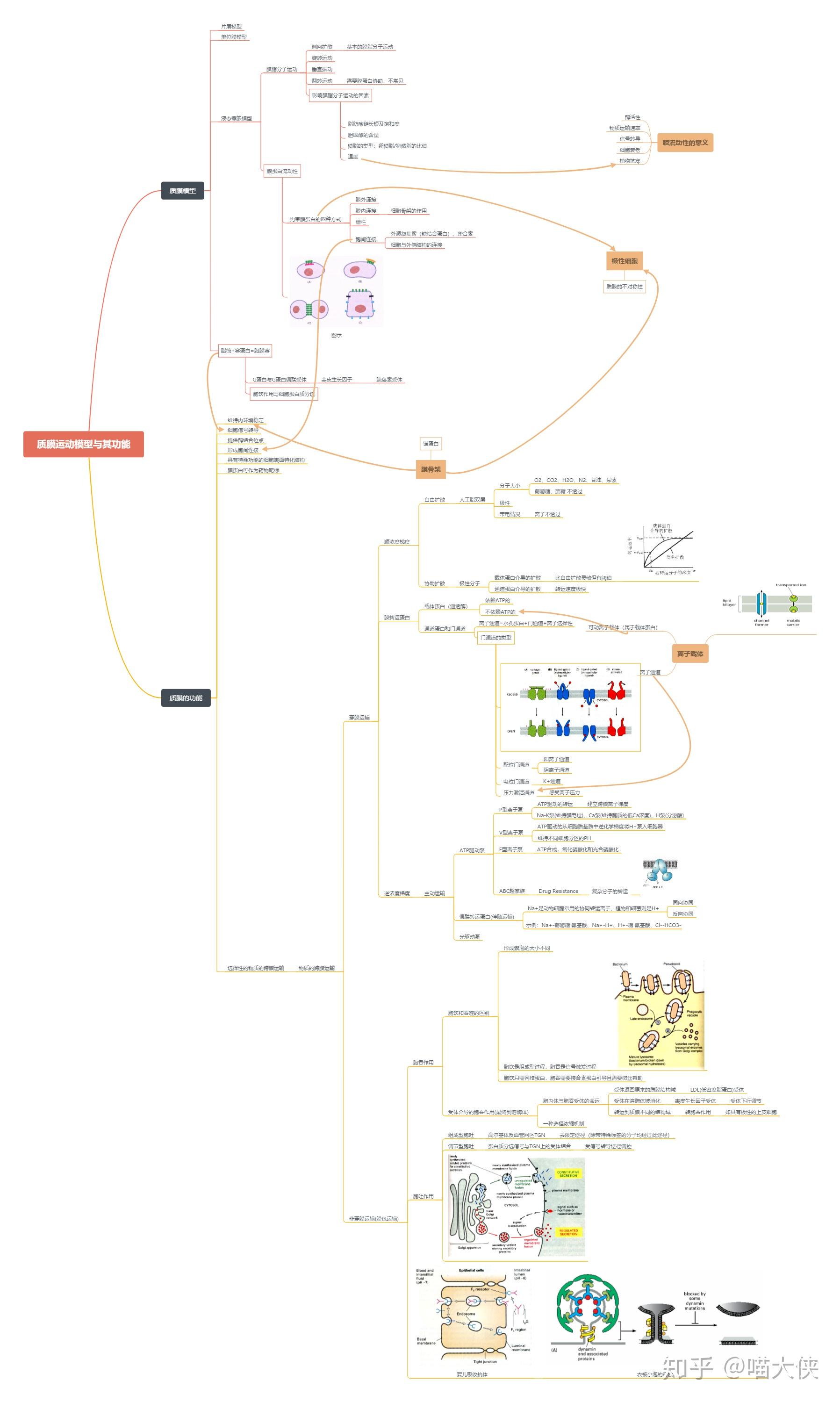
第五章 分子生物学研究方法(上)PPT已看
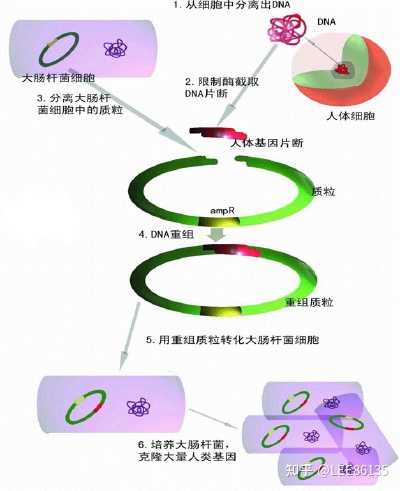
重组DNA技术
DNA基本操作技术
核酸凝胶电泳技术
以琼脂凝胶作为支持物,利用DNA分子在泳动时的电荷效应和分子筛效应,达到分离分子大小不同的DNA的目的。迁移速度与DNA构型有关
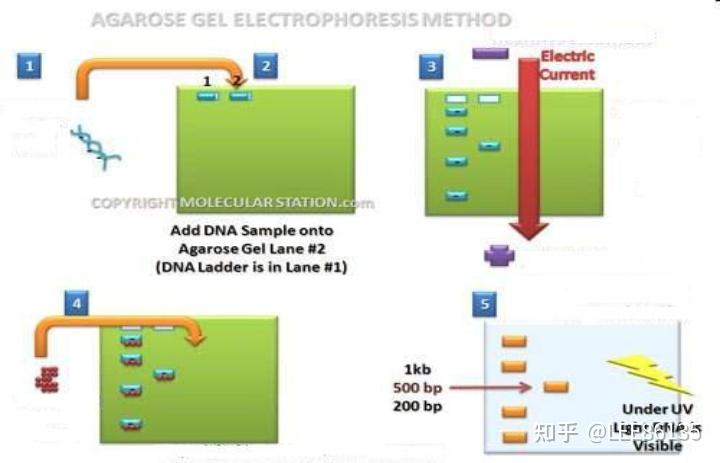
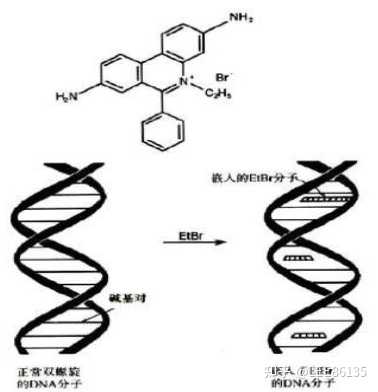
插入溴化乙锭(EB)分子,在紫外光照射下,琼脂糖凝胶电泳中DNA的条带呈现橘黄色荧光,易于鉴定
- 若DNA 太小,用聚丙烯酰胺凝胶电泳技术
- 若DNA 太大,用脉冲电场凝胶电泳(PEGE)技术
通过不断改变电场方向、电流大小、作用时间,使相对MW较大的DNA 更换构型和方位,使其按新方向游动,从而分离。
细菌转化与目标DNA分子的增殖
细菌转化--一种细菌菌株由于捕获了来自供体菌株的DNA而导致性状特征发生遗传改变的过程
感受态细胞(competent cell)——理化方法诱导细胞,使其处于最适摄取和容纳外来DNA的生理状态
$$CaCl_2$$法
不支持在 Docs 外粘贴 block
蓝白斑筛选法:一种基因工程常用的细菌重组子筛选方法
野生型大肠杆菌$$X-gal(5-溴-3-氯-吲哚-β-D-半乳糖苷)\overset{β-D-半乳糖苷}\longrightarrow半乳糖+5-溴-4靛蓝$$
电击法(略)
体外包装法
利用噬菌体颗粒能够有效将 DNA注入到宿主细胞的特点
聚合酶链(PCR)反应技术

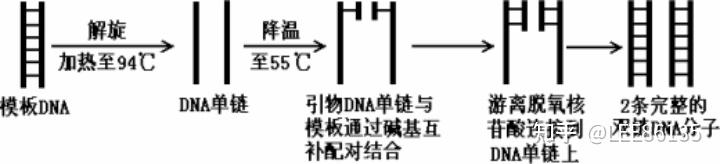
PCR反应的起始材料
- 模板
- 引物
- DNA聚合酶
- dNTP
- $$Mg^{2+}$$
实时定量PCR
利用带荧光检测的PCR仪对整个PCR过程中扩增DNA的累积速率绘制动态变化图,对PCR扩增终产物准确定量
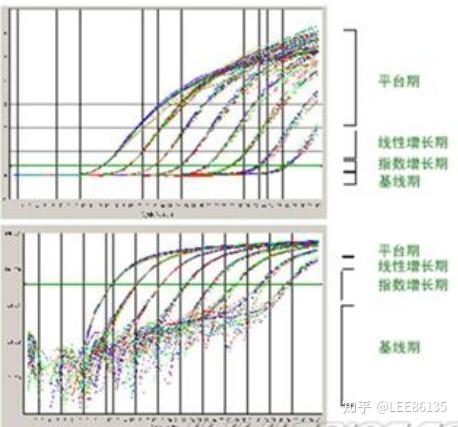
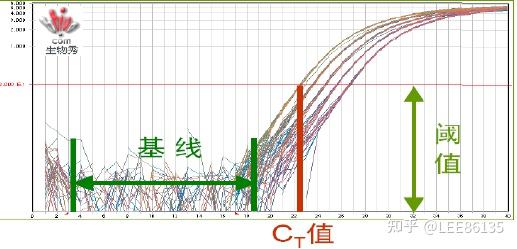
荧光阈值(threshold)的设定
PCR反应的前15个循环的荧光信号作为荧光本底信号,荧光阈值的缺省(默认)设置是3-15个循环的荧光信号的标准偏差的10倍
Ct值(Cycle threshold,循环阈值)
每个反应管内的荧光信号到达设定阈值时所经历的循环数
起始拷贝数越多,Ct值越小。
利用已知起始拷贝数的标准品可作出标准曲线,其中横坐标代表起始拷贝数的对数,纵坐标代Ct值
基因组DNA文库构建
将某生物的全部基因分解成一段一段,每段包含一个或几个基因,再将每个片段通过载体逐一导入到微生物细胞中,所有导入了片段的微生物细胞即为该生物的基因组文库
RNA基本操作技术
总RNA的提取
不能沉淀DNA、蛋白质和多糖
十六烷基三甲基溴化铵,是一种阳离子去污剂
CTAB具有从低离子强度溶液中沉淀核酸与酸性多聚糖的特性。
在高离子强度的溶液中(>0.7M NaCl), CTAB与蛋白质和多聚糖形成复合物,只是不能沉淀核酸。通过有机溶剂抽提,去除蛋白、多糖、酚类等杂质后,加入乙醇沉淀可使核酸分离出来
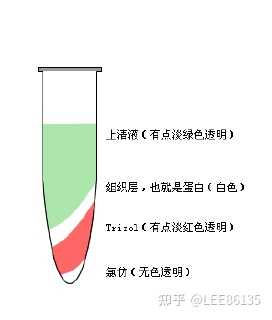
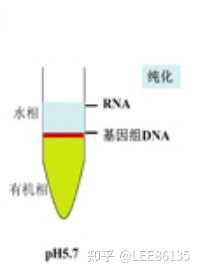
TRIZOL试剂中的主要成分为异硫氰酸胍和苯酚,其中异硫氰酸胍可使RNA与蛋白质分离,并将RNA释放到溶液中。当加入氯仿时,它可抽提酸性的苯酚,而酸性苯酚可促使RNA进入水相。
离心后可形成水相层和有机层, RNA与蛋白质和DNA分离开。
水相层(无色)主要为RNA, 有机层(黄色)主要为DNA和蛋白质。
mRNA的纯化
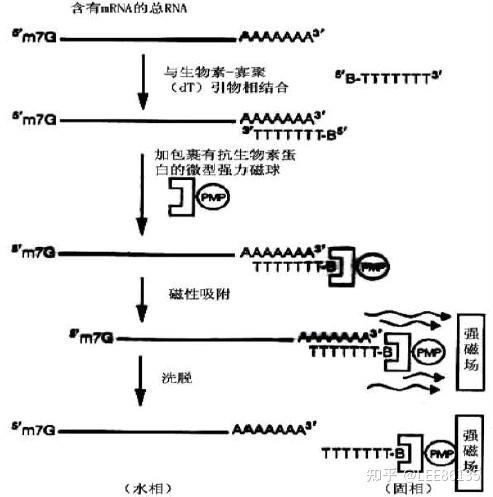
cDNA:与RNA链互补的单链DNA。
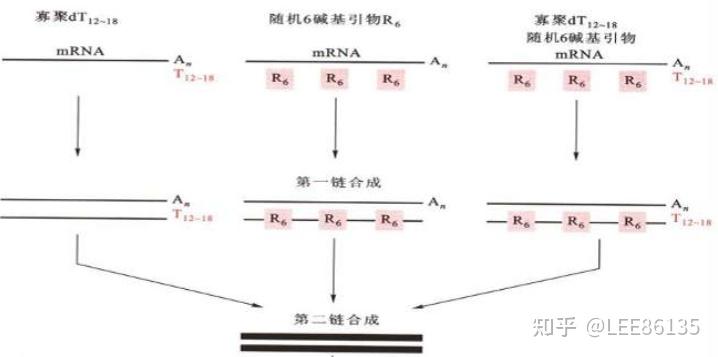
其以RNA为模板,在适当引物的存在下,由RNA与DNA进行一定条件下合成。
cDNA文库--某生物在某发育时期所转录的全部mRNA经反转录形成的cDNA片段,与某种载体连接而形成的克隆的集合
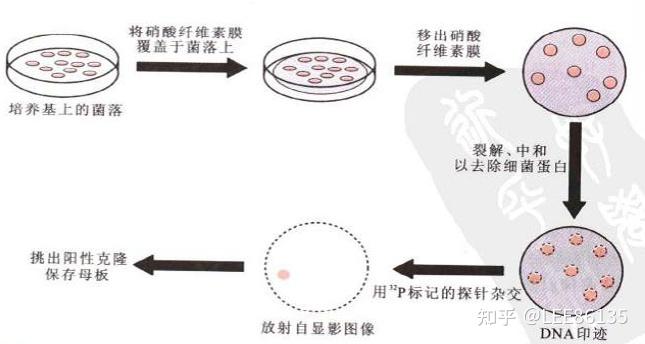
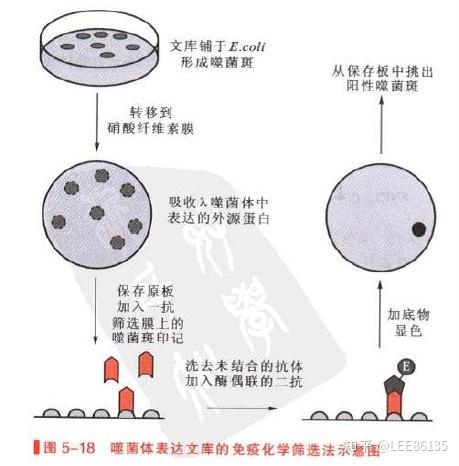
基因文库的筛选
通过某种特殊方法从基因文库中鉴定出含有所需重组DNA分子的特定克隆的过程
SNP
SNP的检测技术
- 基因芯片技术
- Taqman技术
- 分子信标技术
- 焦磷酸测序法
基因克隆技术
cDNA序列可来自序列表达标签(EST)、减法cDNA库、差法显示和基因文库筛选。
- 应用cDNA差示法分析克隆基因
- Gateway大规模克隆技术
- 基因的位图克隆法
通过对许多不同的生态型及大量限制性内切酶和杂交探针的分析,找出与目的基因距离最近的RFLP标记,通过染色体步移技术将位于这两个标记之间的基因片段克隆并分离出来
有具有某种表现型的基因都可得到。首先,通过构建遗传连锁图,将目的基因定位到某染色体的特定位点,并在其两侧确定紧密连锁的RFLP或RAPD分子标记
蛋白质组与蛋白质组学技术
蛋白质组(proteome):本质上指的是在大规模水平上研究蛋白质的特征,包括蛋白质的表达水平,翻译后的修饰,蛋白与蛋白相互作用等,由此获得蛋白质水平上关于疾病发生,细胞代谢等过程的全面认识
蛋白质组学(proteomics):源于蛋白质与基因组学两个词的组合,意指“一种基因组所表达的全套蛋白质”,即包括一种细胞乃至一种生物所表达的全部蛋白质
蛋白质技术
- 双向电泳技术(分离技术)
- 蛋白质印迹法(检测技术)
- 蛋白质的质谱分析技术(鉴定技术)
双向电泳技术(2-D)
第1向基于蛋白质的等电点不同用等电聚焦分离,具有相同等电点的蛋白质无论其分子大小,在电场的作用下都会用聚焦在某一特定位置即等电点处
第2向按分子量的不同用SDS-PAGE分离,把复杂蛋白混合物中的蛋白质在二维平面上分开
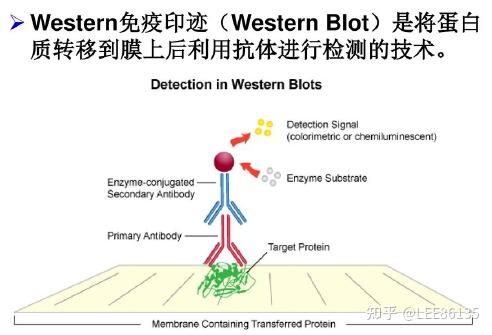
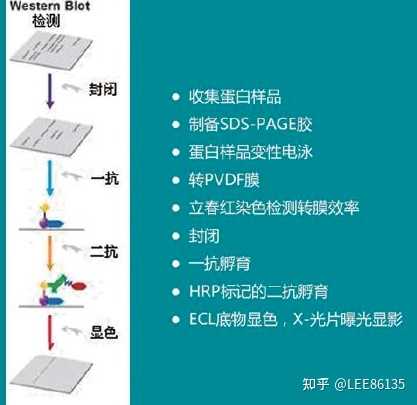
蛋白质印迹法/免疫印迹法
基本原理:通过特异性抗体对凝胶电泳处理过的细胞或生物组织样品进行着色。通过分析着色的位置和着色深度获得特定蛋白质在所分析的细胞或组织中表达情况的信息
蛋白质的质谱分析技术
原理:蛋白质经过蛋白酶的酶切消化后成肽段混合物,在质谱仪中电离形成带电离子,质谱分析器的电场、磁场将肽段离子分离开来,经过检测器收集分离的离子,确定每个离子的质核比值(M/Z)
经过质量分析器可分析出每个肽段的M/Z,得到蛋白质的一级质谱峰图。
离子选择装置自动选取强度较大肽段离子进行二级质谱分析,输出所选肽段的二级质谱峰图,
通过和理论上蛋白质经过胰蛋白酶消化后产生的一级质谱峰图和二级质谱峰图进行比对而鉴定蛋白质
第六章 分子生物学研究方法(下)PPT已看
无法复制加载中的内容
上面的看书理解一下
转录组测序分析和RNAseq
基于传统的Sanger测序法对转录组进行研究的方法主要包括
- 表达序列标签(EST)测序技术
- 基因表达系列分析技术(SAGE)
RNA-seq--转录组测序技术即把mRNA、smallRNA、lncRNA等,用高通量测序技术进行测序分析,反映表达水平
原位杂交技术
用标记的核酸探针,经放射自显影或非放射检测体系,在组织、细胞、间期核及染色体上对核酸进行定位和相对定量研究的一种手段
荧光原位杂交(FISH)首先对寡核苷酸探针做特殊的修饰和标记,然后用原位杂交法与靶染色体或DNA上特定的序列结合,再通过与荧光素分子相耦联的单克隆抗体,来确定该DNA序列在染色体上的位置
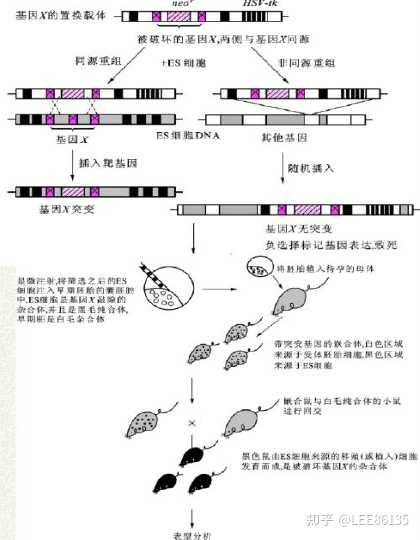
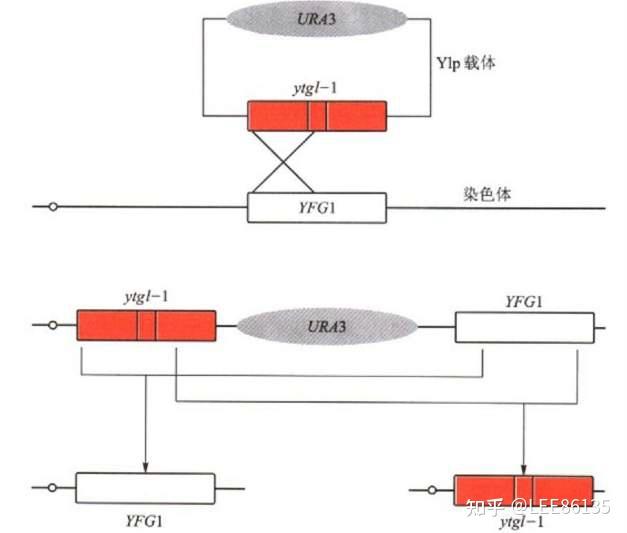
基因敲除技术
通过一定的途径使机体特定的基因失活或缺失的技术
通过同源重组法完全消除细胞或者动物个体中的靶基因活性。
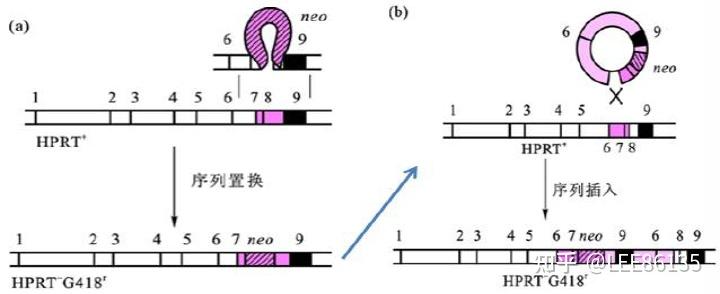
取代型a 插入型b
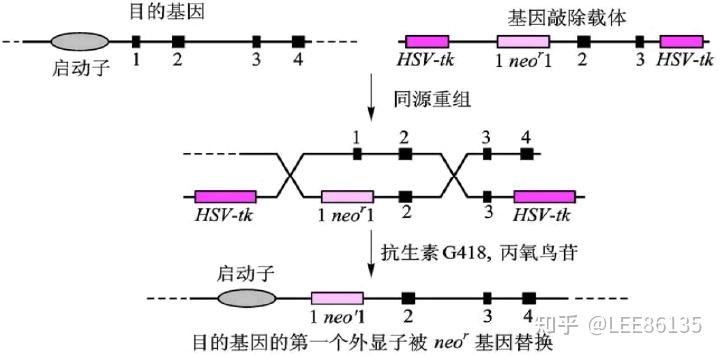
正负筛选法(PNS法)筛选已发生同源重组的细胞示意图(HSV-tk:负向选择基因)
Cre-LoxP重组酶系统
Cre重组酶是一种位点特异性重组酶,能介导两个LoxP位点(序列)之间的特异性重组,使LoxP位点间的基因序列被删除或重组
LoxP(locus of X(cross)-overinP1)序列:来源于P1噬菌体,是由两个13bp反向重复序列和中间间隔的8bp序列共同组成,8bp的间隔序列同时也确定了LoxP的方向
- 在胚胎干细胞的基因组中引入LoxP序列,这一步可以通过打靶载体的设计和对同源重组子的筛选来实现
- 通过Cre介导的重组来实现靶基因的遗传修饰或改变
高等动物基因敲除技术
技术路线主要包括构建重组基因载体,用电穿孔、显微注射等方法 把重组DNA导入胚胎干细胞纯系中,使外源DNA与胚胎干细胞基因组中相应部分发生同源重组,将重组载体中的DNA序列整合到内源基因组中并得以表达
胚胎干细胞(ES细胞)分离和体外培养的成功奠定了哺乳动物基因敲除的技术基础
植物基因敲除技术
T-DNA插入失活技术是目前在植物中使用最为广泛的基因敲除手段
利用根癌农杆菌T-DNA介导转化,将带有报告基因的DNA序列整合到基因组DNA上,如果这段DNA插入到目的基因内部或附近,就会影响该基因的表达,从而使该基因“失活”
蛋白质及RNA相互作用技术
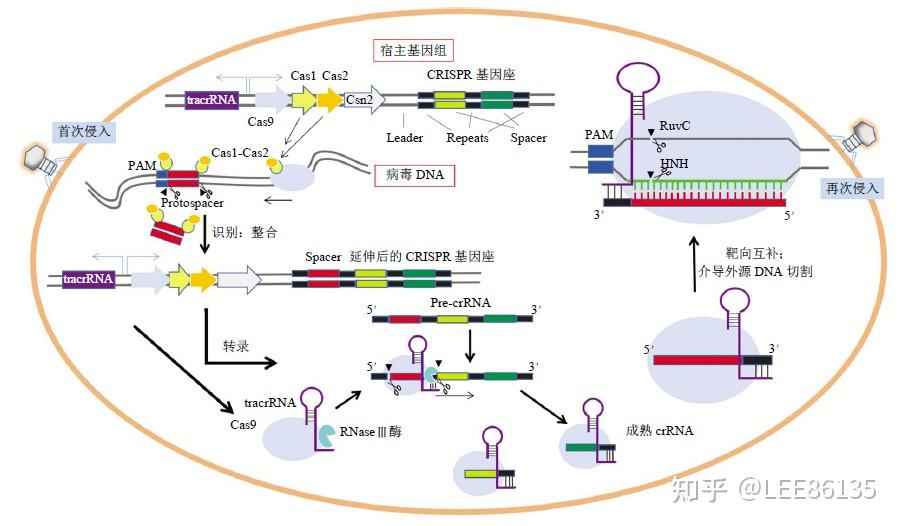
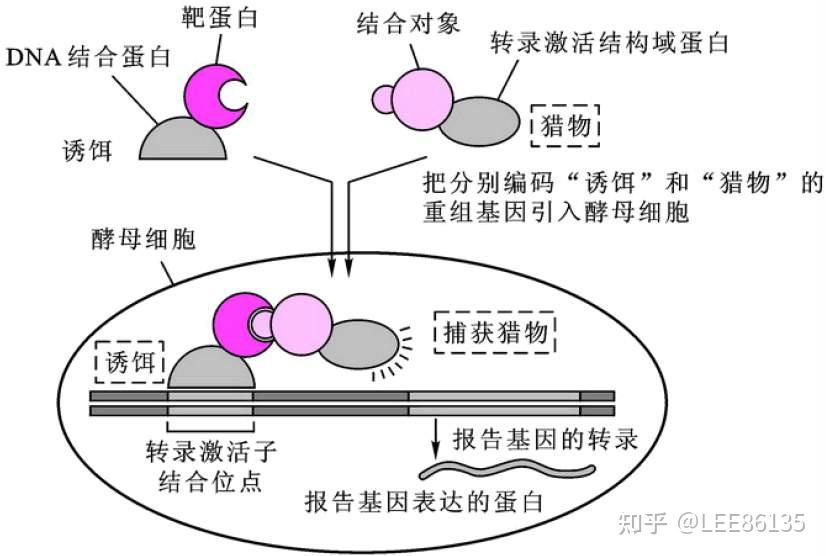
- 酵母单杂交系统。真核生物转录调控因子具有组件式结构特征,这些蛋白往往由两个或两个以上相互独立的结构域,其中 DNA结合结构域(BD) 和转录激活结构域(AD)是转录激活因子发挥功能所必需的。研究DNA-蛋白质之间相互作用
- 酵母双杂交系统。研究蛋白质-蛋白质之间相互作用
- 体外蛋白质相互作用技术:
- 1.Far Western印迹技术
- 2.GST融合蛋白沉降技术
- 3.蛋白质芯片技术
- 4.等离子表面共振技术
- 5.免疫共沉淀技术
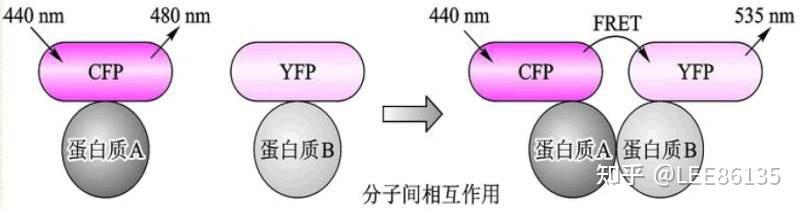
- 细胞内蛋白质相互作用研究—荧光共振能量转移法(FRET)当供体荧光分子的发射光谱与受体荧光分子的吸收光谱重叠,并且两个分子的距离在10nm范围以内时,就会发生一种非放射性的能量转移,即FRET现象,使得供体的荧光强度比它单独存在时要低的多(荧光猝灭),而受体发射的荧光却大大增强(敏化荧光)
通过定位重组系统实现特定时间和空间的基因敲除
噬菌体(黏性与非黏性)会出原题
RNAi技术及其应用
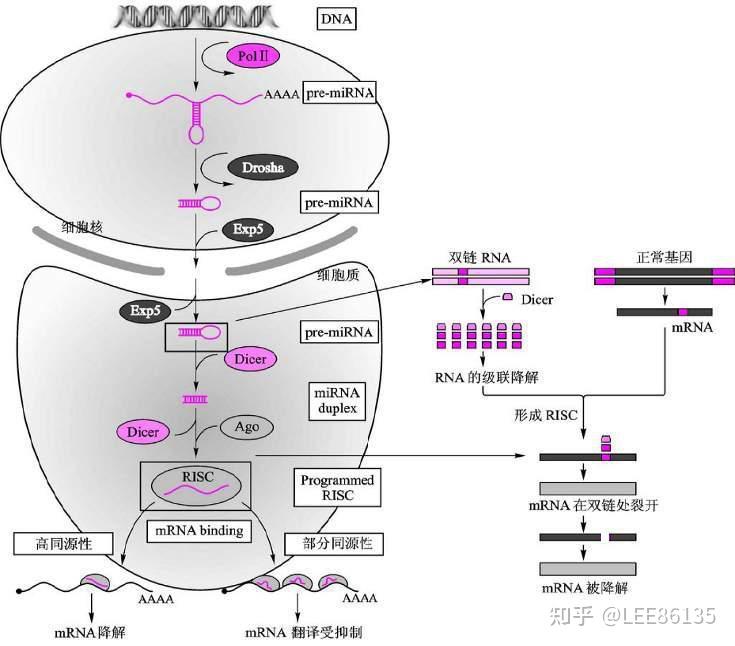
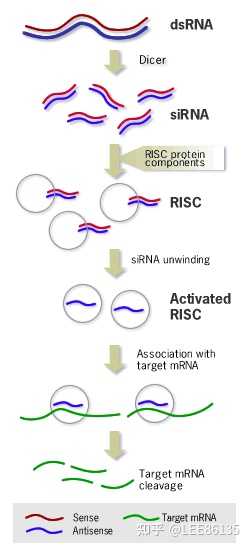
sRNA或pre-miRNA被导入细胞后,能够被一种特异的核酸内切酶(Dicer)识别并切割成为小片段siRNA
这些片段siRNA在RNA解旋酶的作用下解链。继之反义链在与体内一些酶结合形成RNA诱导的沉默复合物(RISC)
RISC与mRNA同源区进行特异性结合,若RISC与mRNA的结合位点完全配对,则降解mRNA;若RISC与mRNA的结合位点不完全配对,则抑制mRNA的翻译
基因芯片及数据分析
基因芯片,又称DNA微阵列技术是能同时监测大量靶基因表达的实验手段,从而迅速准确地在基因组水平上阐述不同生物组织或细胞中各种转录本的变化规律
利用酵母鉴定靶基因功能
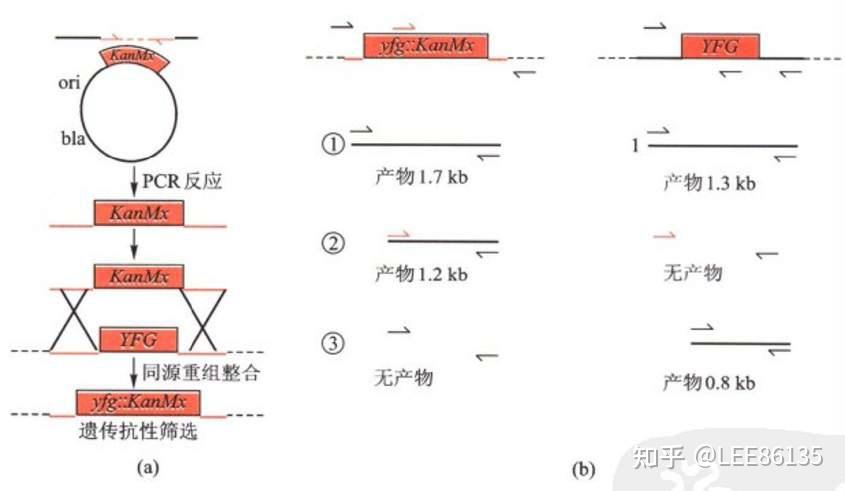
“一步置换法”制备酵母缺失突变体的流程图 “两步置换法”
其他分子生物学技术
凝胶滞缓实验(EMSA)
凝胶滞缓试验,又称DNA迁移率变动试验,是用于体外研究DNA与蛋白质相互作用的一种特殊的凝胶电泳技术
在凝胶电泳中,由于电场的作用,裸露的DNA朝正电极移动的距离与其分子量的对数成反比。如果此时DNA分子与某种蛋白质相结合,那么,由于分子量增大,它在凝胶中的迁移作用便会受到阻滞,在特定电压和时间内朝正电极移动的距离也就相应缩短了
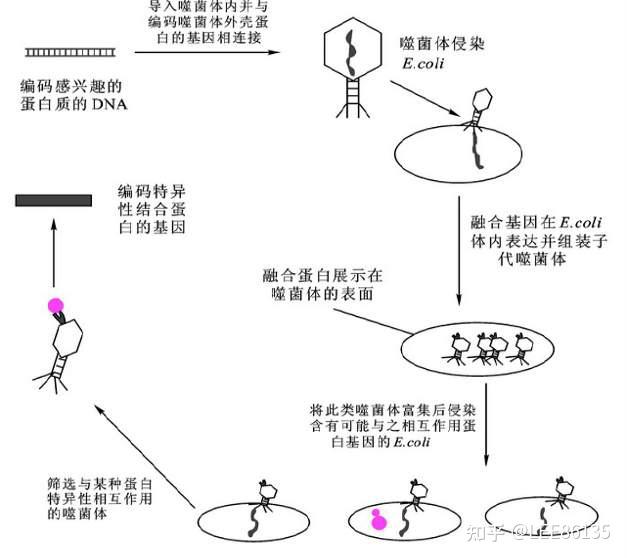
噬菌体展示技术
噬菌体可被分为溶菌周期和溶源周期两种不同类型
烈性噬菌体(只有溶菌周期)、温和噬菌体(有溶源周期)
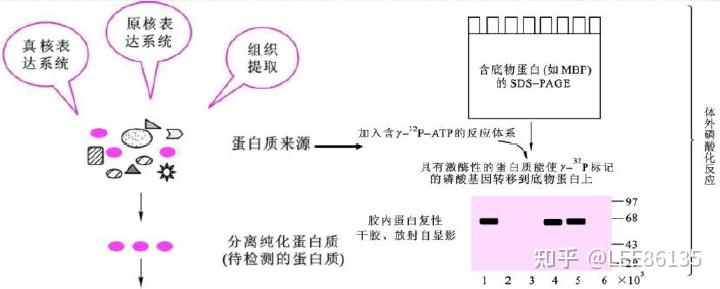
蛋白质磷酸化分析技术
磷酸激酶--促使底物蛋白质磷酸化
蛋白质磷酸脂酶--催化蛋白质发生去磷酸化
一般采用双向电泳及质谱分析法检测底物蛋白磷酸化
蛋白免疫印迹实验
全基因组关联分析--挖掘型状变异的相关基因
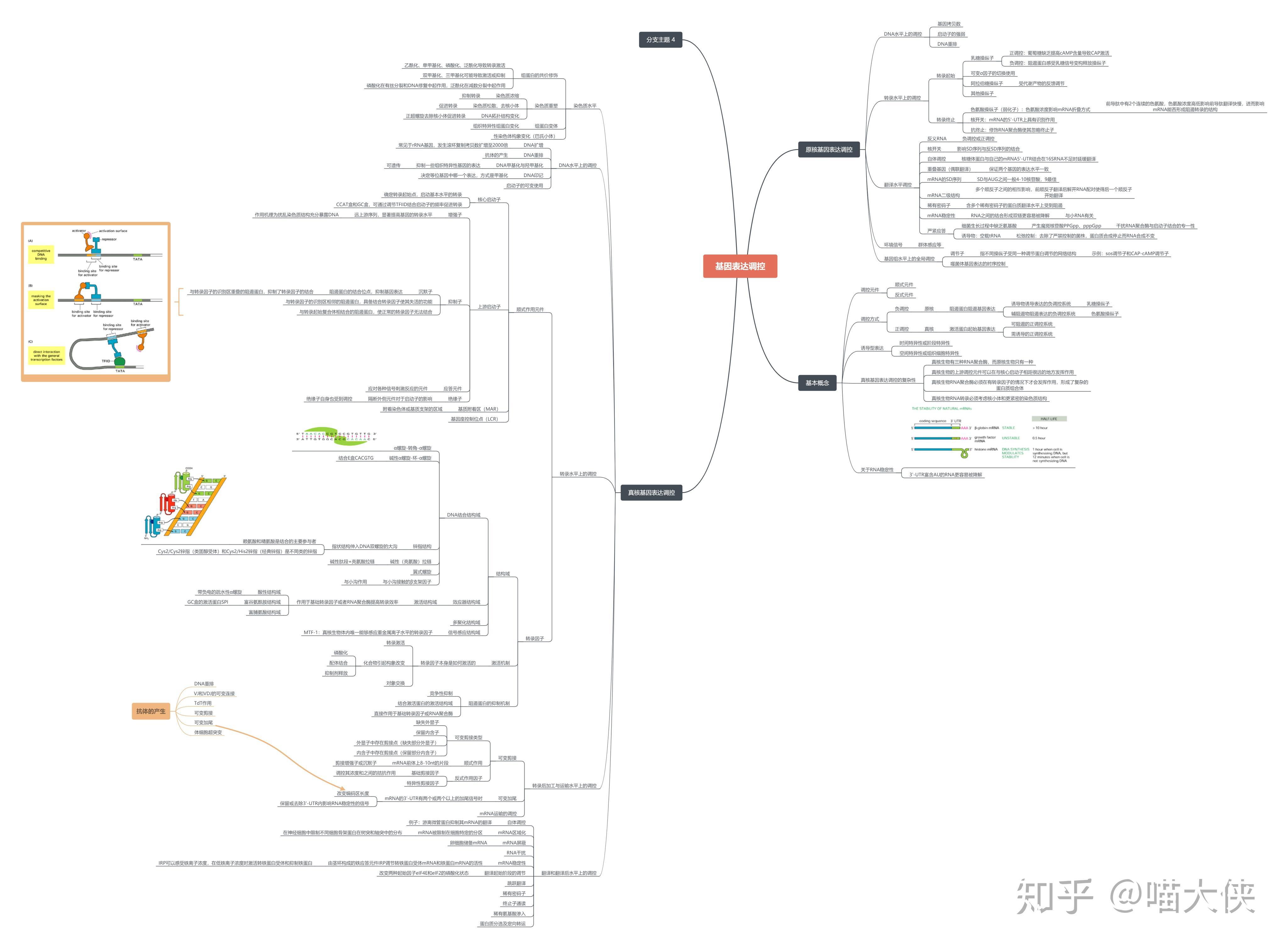
第七章 基因的表达与调控 PPT已看
原核基因表达调控模式
重点是乳糖操纵子、色氨酸操纵子,其他地方从书里出一点题
基因表达--基因转录及翻译,rRNA与tRNA的合成也属于
无法复制加载中的内容
原核基因调控
无法复制加载中的内容
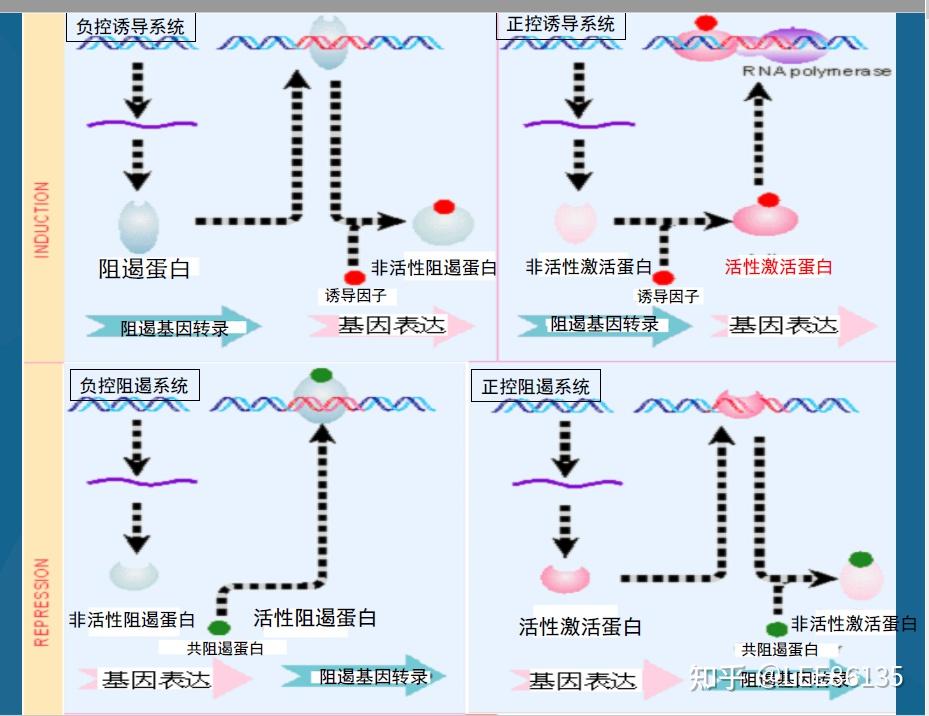
- 适应性调节--为适应环境的变化而做出的调节
- 转录水平的调节
- 以操纵子为单位,有正、负调控两种机制
可诱导调节
可阻遏调节
弱化子
当操纵子被阻遏,RNA合成被终止时,起终止转录信号作用的那段核苷酸。
有特殊负载的氨酰-tRNA的浓度起信号作用。属于转录调节中的微调整
降解物对基因活性的调节
葡萄糖效应:只有体系中无葡萄糖,原核生物才会开始利用乳糖
降解物抑制作用是通过提高转录强度来调节基因表达的,是一种积极的调节方式
细菌的应急反应
信号--鸟苷四磷酸、鸟苷五磷酸
产生这两种物质的诱导物是空载tRNA
当AA饥饿时,存在大量空载tRNA,激活焦磷酸转移酶,使鸟苷四磷酸大量合成。鸟苷四磷酸关闭许多基因,并打开AA合成基因
影响一大批操纵子,属于超级调控因子
操纵子:启动基因、操纵基因和一系列紧密连锁的结构基因的总称
乳糖操纵子
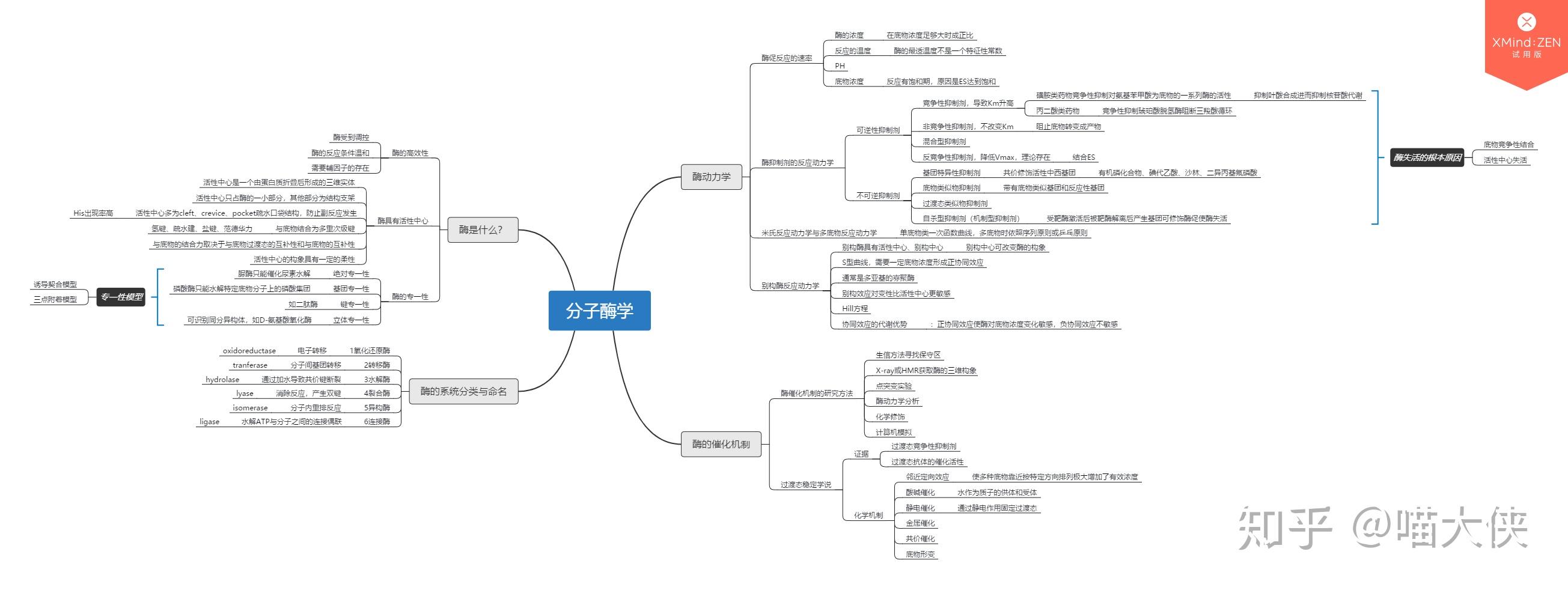
酶的诱导--lac体系受调控的证据
当有乳糖供应时,在无葡萄糖培养基中生长的lac细菌中将同时合成β-半乳糖苷透过酶、β-半乳糖苷酶--表明乳糖能激发lac mRNA合成
安慰诱导物--促使细菌产生酶而本身不被分解的物质
异丙基巯基半乳糖苷IPTG 巯甲基半乳糖苷TMG O-硝基半乳糖苷ONPG
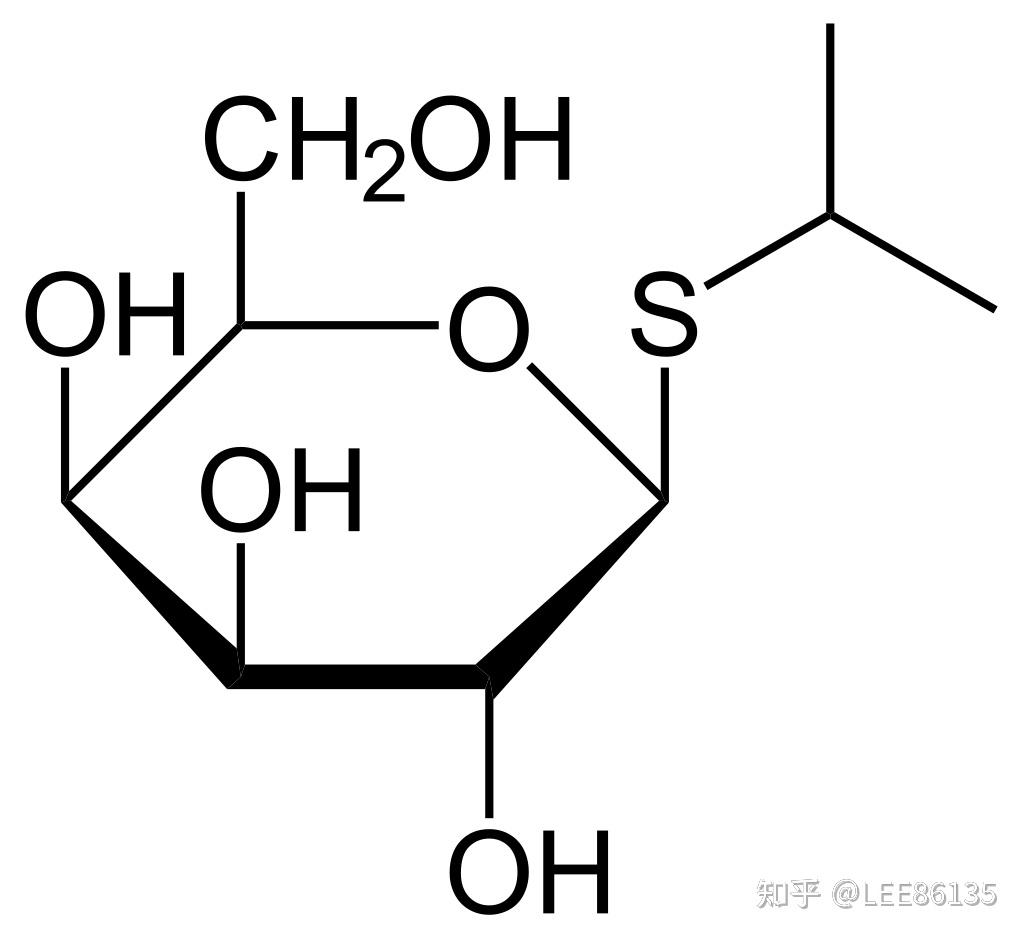
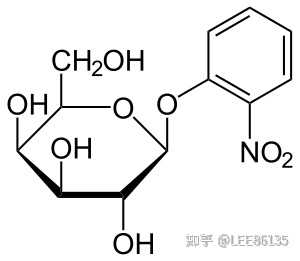
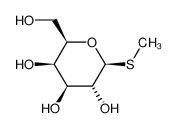
酶合成不是由前体转化,而是由诱导物诱导新合成--$$^{35}S$$同位素示踪实验
操纵子模型
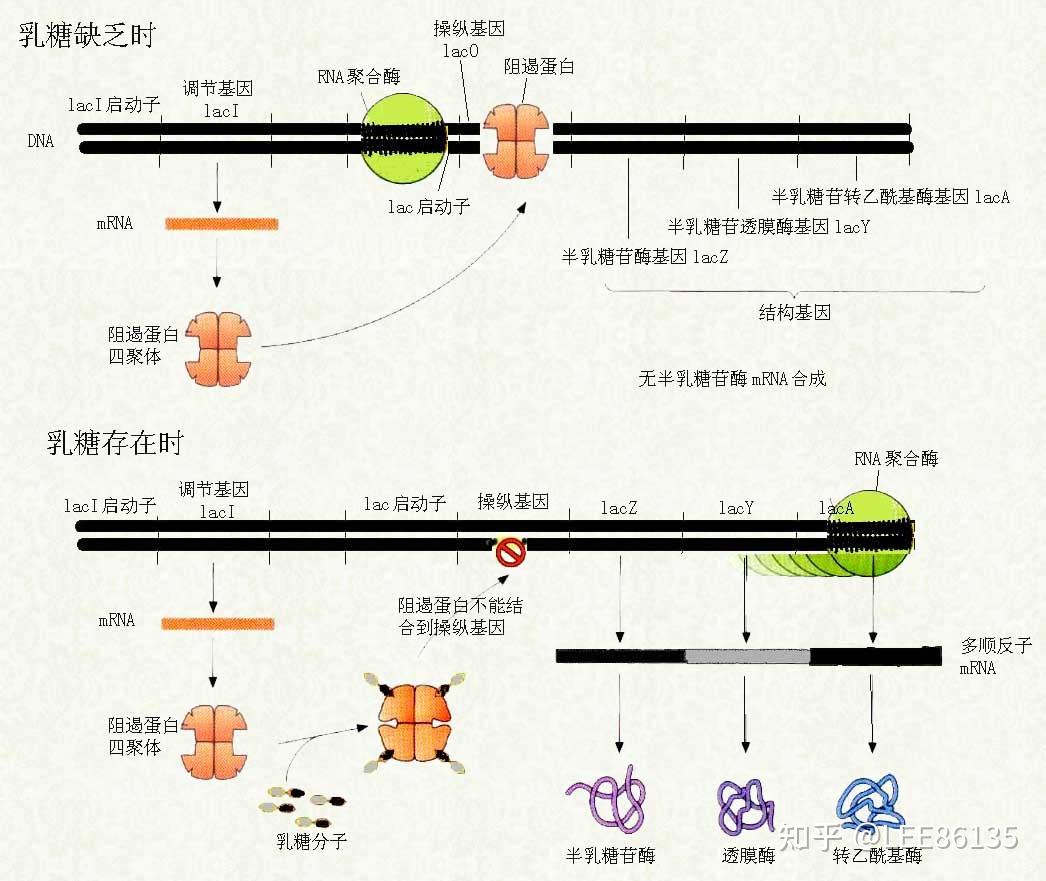
操纵子影响因子
操纵子理论不能解释的问题
- 诱导物需要穿过细胞膜才能与阻遏物结合,转运诱导物需要透过酶,透过酶需要合成。第一个诱导物如何进入细胞?
一些透过酶可在无诱导物的情况下合成
- 真正的诱导物是乳糖在β-半乳糖甘酶催化下形成的异构乳糖,第一个β-半乳糖甘酶如何合成的?
非诱导状态下有少量lac mRNA 合成
无法复制加载中的内容
乳糖操纵子阻遏物mRNA是由弱启动子控制下永久型合成的
当阻遏基因由弱启动子突变成启动子,细胞内就不能产生足够的诱导物来克服阻遏状态,整个乳糖操纵子在这些突变体中不可诱导
若培养基中存在葡萄糖,则乳糖操纵子处于阻遏状态,不能被诱导
cAMP形成:$$ATP\overset{腺苷酸环化酶}\longrightarrow cAMP(环腺苷酸)$$
代谢激活蛋白CRP(又称CAP)由Crp基因编码,它可与cAMP形成复合物,是激活乳糖操纵子的正调控因子
cAMP-CRP不但能与DNA相结合,造成双螺旋弯曲,易于形成三元转录起始复合物,还能直接影响RNA聚合酶的活性
大肠杆菌中,cAMP浓度受葡萄糖代谢调节
- 缺乏碳源,cAMP浓度高,CAP增加,促进转录
- 有葡萄糖,cAMP浓度低,CAP减少,不促进转录
- 只有甘油或乳糖等不进行糖酵解途径的碳源,cAMP浓度高,CAP增加,促进转录
A结构基因及其生理功能
无法复制加载中的内容
A结构基因的作用是抑制β-半乳糖苷酶产物的有害产物在细胞内积累
翻译水平的调节导致不同lac基因产物酶在数量上的差异
β-半乳糖苷酶:透过酶:乙酰基转移酶=1:0.5:0.2
- Lac mRNA可能与翻译过程中的核糖体相脱离,从而终止蛋白质链的翻译
- 在lac mRNA分子内部,A基因比Z基因更容易受到内切酶作用发生降解
操纵子的融合与基因工程(基因融合技术)
lac操纵子与pur操纵子偶联
色氨酸操纵子
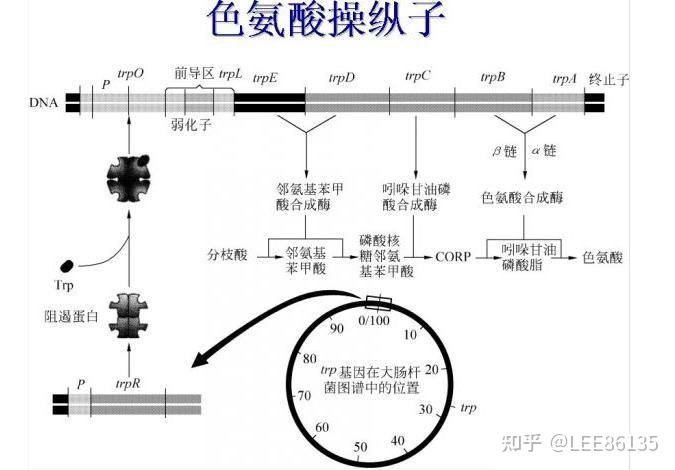
- trpR和trp ABCDE不连锁
- 操纵基因在启动子内
- 有衰减子(又称弱化子)
- 启动子和结构基因不能直接相连,二者被前导序列隔开
负控阻遏系统
trpR基因突变常引起trp mRNA的永久性合成,该基因产物被称为辅阻遏蛋白
该阻遏系统中的效应物分子是色氨酸trp
当培养基中色氨酸含量较高时,色氨酸与游离阻遏蛋白结合,并使之与操纵区DNA紧密结合
当培养基中色氨酸含量较低时,游离阻遏蛋白失去辅阻遏物并从操纵区上解离,trp操纵子去阻遏
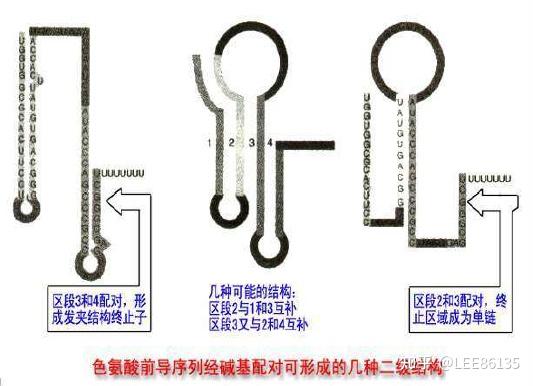
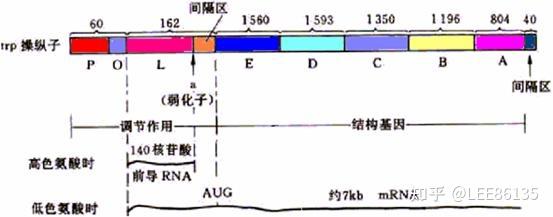
弱化机制
弱化子--DNA中可导致转录过早终止的一段核苷酸序列,位于前导序列L中
该区mRNA通过自我配对可形成茎环结构,有典型终止子的特点
前导肽--由前导序列翻译出来的一段肽链
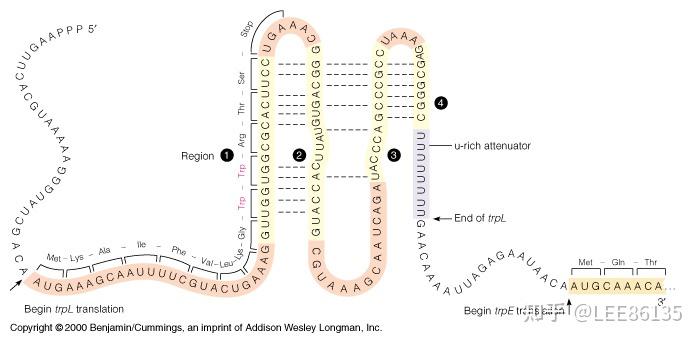
形成发夹结构能力强弱:1+2>2+3>3+4
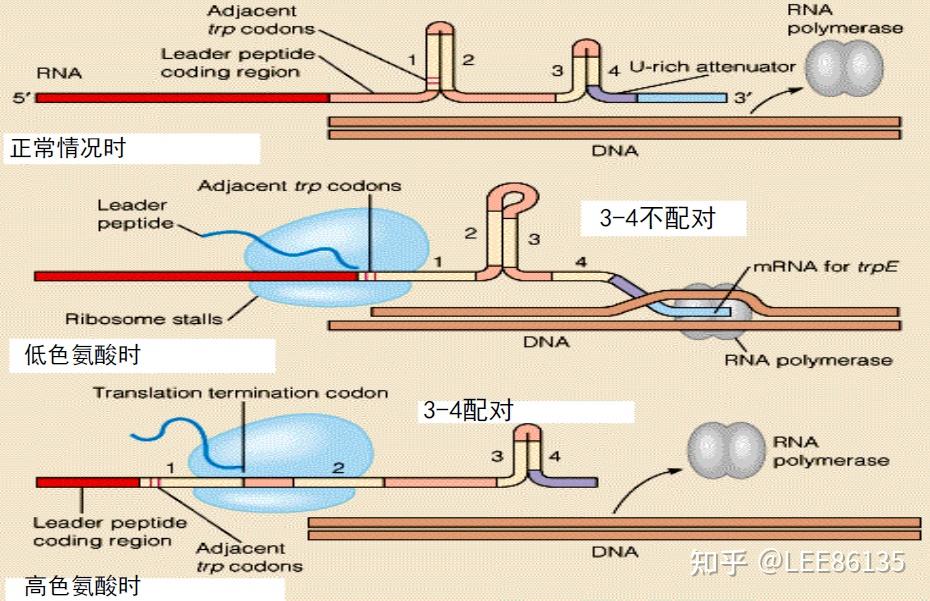
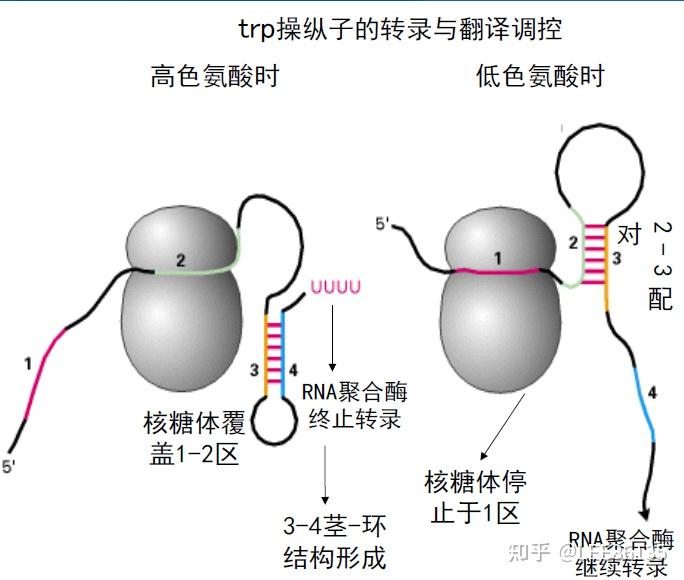
阻遏作用只能使转录不起始;弱化作用可使转录中途终止
阻遏作用信号是细胞内色氨酸含量;弱化作用信号是细胞内载有色氨酸的tRNA的含量。
色氨酸浓度高时,序列3+4形成衰减子结构,不转录后续基因
色胺酸浓度低时,序列2+3结合,不形成衰减子,转录后续基因
半乳糖gal操纵子
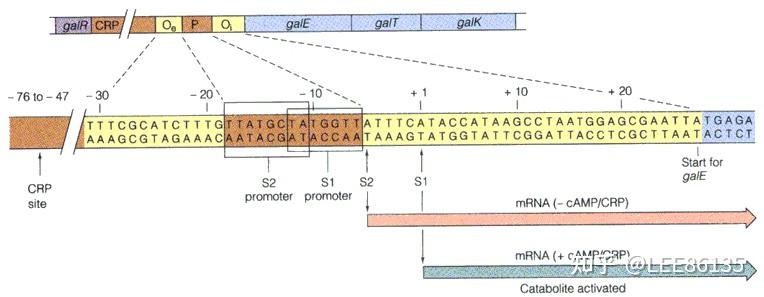
结构基因:galE编码异构酶;galT编码乳糖-磷酸尿嘧啶核苷酸转移酶;galK编码半乳糖激酶
gal操纵子有两个启动子;有两个O区,分别在P区上游和结构基因galE内部
阿拉伯糖ara操纵子
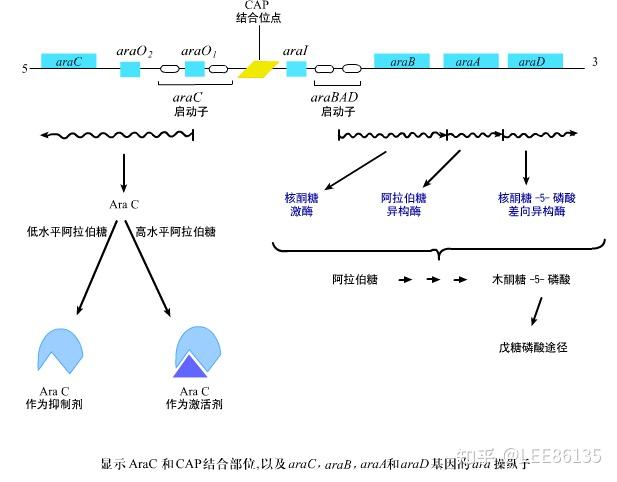
araC基因产物调控三个操纵子编码araBAD、araE、araF
araC表达受到araC自身调控
araC即是araC操纵子的正调节蛋白(需cAMP-CRP的共同参与起始转录),又是其负调节蛋白。这种双重功能通过araC蛋白的两种异构体实现(Pi和Pr)

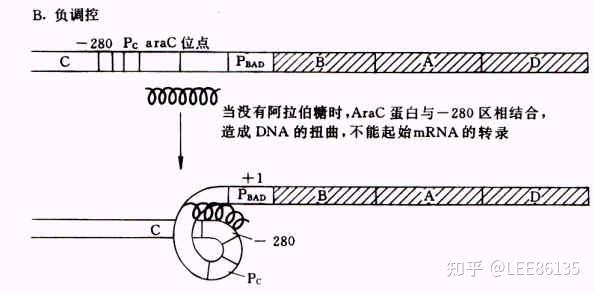
Pr起阻遏作用;Pi起诱导作用
无法复制加载中的内容
阻遏蛋白LexA的降解细菌中的SOS应答
正常情况下,LexA蛋白阻遏SOS基因的表达
紫外线可激活RecA,降解LexA蛋白,表达SOS基因修复DNA
二组分调控系统和信号转导
传感蛋白(细胞膜)
无法复制加载中的内容
多启动子调控的操纵子
rRNA操纵子
大肠杆菌rRNA操纵子上有两个启动子P1、P2。
- 营养充沛时,由强启动子P1起始的转录产物比P2起始的转录产物高3~5倍
- 营养匮乏时,P1的作用被抑制,但P2仍可以发挥作用
核糖体蛋白SI操纵子
强启动子--P1、P2,平时启动基因表达,合成SI蛋白
弱启动子--P3、P4,紧急情况下,P1、P2启动子受ppGpp抑制,由P3、P4起始合成的SI蛋白维持生命最低需要
DNA Q蛋白操纵子
DNA Q蛋白是DNA聚合酶全酶的亚基之一,主要功能是校正DNA复制中出现的错误
当RNA聚合酶活性低时,操纵子由弱启动子P2控制
当RNA聚合酶活性高时,操纵子由强启动子P1控制
固氮基因调控
固氮酶
无法复制加载中的内容
固氮酶促反应
$$N_2+16MgATP+8e^-+8H^+\longrightarrow 2NH_3+16MgADP+16Pi+H_2$$

转录水平上的其他调控方式
σ因子的调节作用
热休克蛋白HSP
由$$\sigma^{32}$$参与构成的RNA聚合酶与热休克应答基因启动子结合,诱导产生大量热休克蛋白,适应环境需要
枯草芽孢杆菌芽孢形成
有序的σ因子替换,RNA聚合酶识别不同基因的启动子,使芽孢形成有关基因有序表达
组蛋白类似蛋白调节
组蛋白类似蛋白--组蛋白中非特异性DNA结合蛋白,维持DNA的高级结构
转录后调控
翻译起始的调控
RBS核糖体结合位点--mRNA链上起始密码子AUG上游的一段非翻译区
RBS的结合强度取决于SD序列的结构与起始密码子AUG之间的距离
稀有密码子对翻译的影响
细胞内对应于稀有密码子的tRNA较少,高频使用这些密码子的翻译易受阻
重叠基因对翻译起始的调控
trpE的终止密码子与trpD起始密码子重叠
翻译终止时核糖体立即处在起始环境中,重叠密码子保证了同一核糖体对连续基因进行翻译的机制

翻译阻遏对起始的调控
Qβ噬菌体的复制酶,可结合于SD序列,阻断核糖体在mRNA上的移动,抑制了翻译
魔斑核苷酸水平对翻译的影响
紧缩调节--与培养基环境变化相应而发生的暂时性细胞繁殖抑制,及促进适应新环境的调节机能等
RC基因--RNA control,并分为RC约束型与RC松弛型,又称为rel+、rel-型
在氨基酸缺乏时,rel菌能大量合成两种特殊核苷酸:鸟苷四磷酸ppGpp;鸟苷五磷酸pppGpp
第八章 基因的表达与调控(下)PPT已看
断裂结构
结构特点要考
外显子、内含子
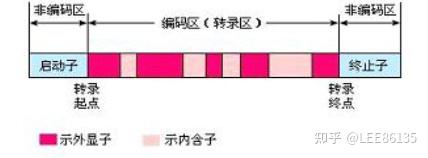
真核生物细胞基因由编码区与非编码区两部分组成。
非编码区上有调控序列。
编码区是不连续的,能够编码蛋白的序列被不编码蛋白的序列隔开,成为一种断裂的形式。
编码区叫外显子,非编码区叫内含子。
连接区
外显子-内含子连接区的高度保守性和特异性碱基序列
几乎每个内含子5’端起始的两个碱基都是GU(T),而3’端最后两个碱基总是AG,由于这两个碱基的高度保守性和广泛性,有人把它称为GT-AG法则--要考
真核基因的原始转录产物可通过不同的剪接产生不同的mRNA,翻译成不同的蛋白质。
组成型剪接--一个基因的转录产物通过组成型剪接只能产生一种成熟的mRNA,编码一个多肽。如肌红蛋白重链基因虽有41个外显子,却能精确地剪接成一个成熟的mRNA。
选择性剪接--同一基因的转录产物由于不同的剪接方式形成不同mRNA的过程。
基因家族、基因簇
基因家族:真核细胞中相关基因按功能成套组合
- 简单多基因家族--基因以串联方式相连
- 复杂多基因家族--基因家族之间由间隔序列隔开,并作为独立的转录单位
- 发育调控的复杂多基因家族--血红蛋白在生物个体发育的不同阶段出现几种不同形式的α和β亚基
真核基因表达的方式和特点
原核细胞——主要是通过转录调控,以开启或关闭某些基因的表达来适应环境条件。环境因素对调控起决定性的作用
真核细胞——基因表达调控最明显的特征是在特定时间,特定的细胞中特定的基因被激活,实现“预定”的、有序的、不可逆转的分化、发育,并使生物的组织和器官保持正常功能
名词性解释
下面的思维导图
无法复制加载中的内容
问答题--真核基因表达的转录水平调控
真核基因调控主要在转录水平上进行,受大量特定的顺式作用元件和反式作用因子调控
顺式作用元件:指启动子和基因的调节序列,主要包括启动子、增强子、沉默子等。
无法复制加载中的内容
增强子
指能使与它连锁的基因转录频率明显增加的DNA序列
增强子通常具有下列特性:
- 增强效应十分明显。
- 增强效应与其位置和取向无关。
- 大多为重复序列(50bp)。
- 其增强效应有严密的组织和细胞特异性。
- 无基因专一性,可在不同的基因组合上表现增强效应。
- 许多增强子还受外部信号的调控
反式作用因子:指能直接或间接地识别或结合在各类顺式作用元件上调控基因表达的蛋白质或RNA。多为转录因子
被广泛研究的转录因子主要有:TFIID--识别TATA区;CTF--识别CAAT区;SP1--识别GGGCGG;HSF--识别热激蛋白启动区
DNA识别域或结合域的五种结构
- 螺旋-转折-螺旋结构 (HTH)
- 锌指结构(zinc finger)
- 碱性-亮氨酸拉链(bZIP结构)
- 碱性-螺旋-环-螺旋(bHLH结构)
在免疫球蛋白κ轻链基因的增强子结合蛋白E12与E47中,羧基端100~200个氨基酸残基可形成两个两性α螺旋,被非螺旋的环状结构所隔开,蛋白质的氨基端则是碱性区,其DNA结合特性与亮氨酸拉链类蛋白相似
同源域蛋白是指编码60个保守氨基酸序列的DNA片段
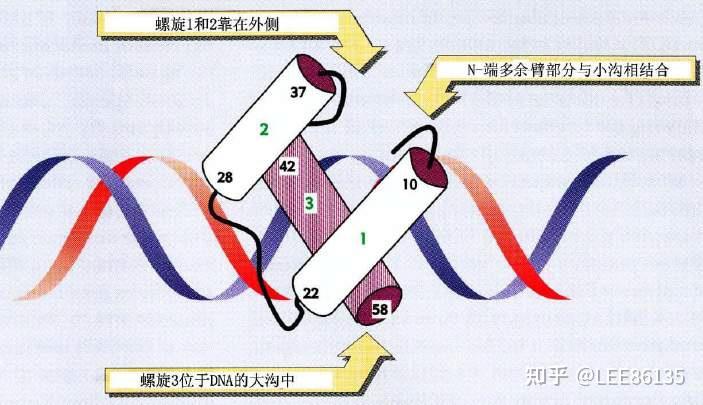
同源域蛋白形成3个α螺旋。C端α螺旋有17个氨基酸,结合DNA大沟,N端臂插入DNA小沟
不同的转录活化域,有下列特征性结构
- 带负电荷的螺旋结构
- 富含谷氨酰胺(Gln)的结构
- 富含脯氨酸(Pro)的结构
真核基因表达的染色体修饰和表观遗传调控
真核生物DNA水平上的基因表达调控
DNA水平的调控是真核生物发育调控的一种形式,包括基因丢失、扩增、重排和移位等。这种调控使基因组发生了改变。
如:一个成熟红细胞能产生大量的可翻译成熟珠蛋白的mRNA,它的前体细胞是不产生珠蛋白的。这种变化是由于基因本身或者是基因的拷贝数发生了永久性变化所调控的
活跃状态的基因更容易被DNA酶I所降解。基因活跃表达时启动区部分序列可能解开成单链,从而不能继续缠绕在核小体上,使启动区DNA“裸露”于组蛋白表面,形成对DNA酶I的超敏感现象
基因扩增是指某些基因的拷贝数专一性大量增加的现象,它使细胞在短期内产生大量的基因产物以满足生长发育的需要,是基因活性调控的一种方式
将一个基因从远离启动子的地方移到较近的位点从而启动转录,被称为基因重排。
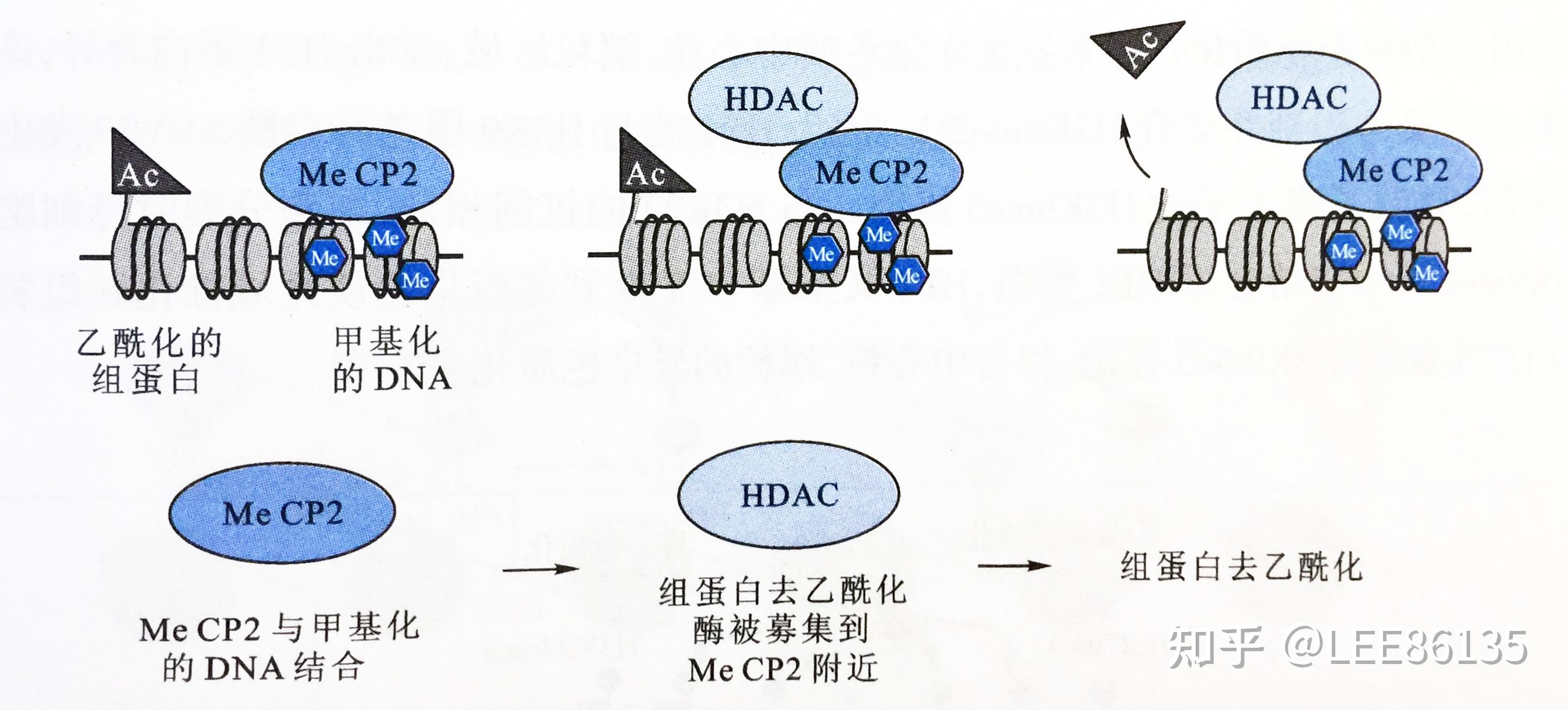
DNA甲基化与基因活性调控
DNA甲基化能引起染色质结构、DNA构象、DNA稳定性、DNA与蛋白相互作用方式的改变,从而控制基因表达
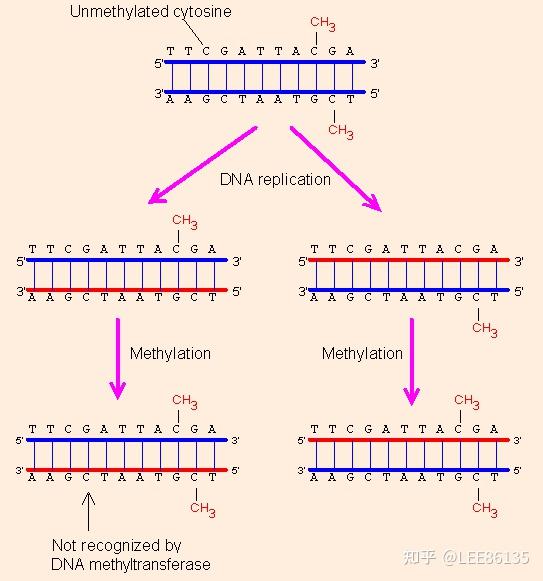
- 日常型甲基转移酶使半甲基化的DNA双链上与胞嘧啶(C)相对应的胞嘧啶甲基化
- 从头合成型甲基转移酶催化未甲基化的CpG成mCpG,不需母链指导
甲基化引起DNA构型发生变化:B-DNA→Z-DNA,不利于转录起始
对于弱启动子来说,稀少的甲基化就能使其完全失去转录活性
当这一类启动子被增强时,即使不去甲基化也可以恢复其转录活性。
若进一步提高甲基化密度,会使增强后的启动子恢复不了转录活性。
DNA的甲基化还会提高突变频率
X染色体上有一个重要基因Xist,该基因只在失活的X染色体上表达,而不在活性的X染色体上表达;
活性X染色体上的Xist基因总是甲基化的,而失活X染色体上该位点都是去甲基化的
组蛋白乙酰化对基因表达的影响
蛋白质乙酰化:在乙酰基转移酶的作用下,在蛋白质特定的位置添加乙酰基的过程。蛋白质乙酰化是细胞控制基因表达、蛋白质活性或生理过程的一种机制
组蛋白乙酰化是一可逆的动态过程
组蛋白乙酰化与基因活性的提高有关
机制:组蛋白N端“尾巴”上赖氨酸残基的乙酰化中和了组蛋白尾巴的正电荷,降低了它与DNA的亲和性,导致核小体构象发生有利于转录调节蛋白与染色质相结合的变化,从和提高了基因转录的活性。
组蛋白去乙酰化与基因活性的阻遏有关
组蛋白甲基化对于真核基因表达的调控
H3K9me3标记通常与异染色质化有关。
异染色质化区域,分为两类:
- 非组成型异染色质化: 多发生在不同发育时期一些需要被沉默的基因区域。
- 组成型异染色质化:通常发生在染色质中心粒、端粒区域,常由HP1蛋白介导。
表观修饰的遗传:‘玳瑁猫’常表达染色质修饰的整合作用
可以被甲基化修饰的残基:赖氨酸、精氨酸
组蛋白赖氨酸甲基转移酶HKMT;蛋白精氨酸甲基转移酶PRMT: I型 & II型(催化类型不同)
组蛋白去甲基化酶
基因沉默/RNA沉默
指真核生物中由双链RNA诱导的识别和清除细胞中非正常RNA的一种机制
RNA干扰(RNA interference、RNAi)是指由双链RNA(double-stranded RNA,dsRNA)诱发的、同源mRNA高效特异性降解的技术
由于RNAi技术可以特异性降低或关闭目的基因表达,已被广泛应用于探索基因功能和疾病的基因治疗领域
siRNA
siRNA来源:病毒RNA、环境实验因素引入的外源RNA、以及基因组重复片段、转座子也可能产生的siRNA等。
通常一个长度为21nt的双链小RNA,其中19nt形成配对双链,3’端各有两个不配对核苷酸,5’端为磷酸基团。
一条链为引导链(guide strand)介导mRNA的降解;其中一条链为乘客链(passenger strand),在iRNA形成有功能的复合物前被降解
产生
- Dicer切割
- R2D2的装配
- RISC的装配和成熟
生物学意义
- 在转录水平、转录后水平参与基因的表达调控;
- 维持基因组的稳定;
- 保护基因组免受外源核酸的入侵
miRNA
和siRNA一样装载成RISC后使互补配对的mRNA降解;
microRNA可抑制mRNA的翻译,降低靶基因的蛋白水平,但不影响其mRNA水平。
miRNA-RISC的原理与siRNA相似,但是由此介导的基因沉默需要一些其他功能保守的原件参与
蛋白质磷酸化
受体分子活化细胞功能的途径
- 受体本身或受体结合蛋白具有内源酪氨酸激酶活性,胞内信号通过酪氨酸激酶途径得到传递。
- 配体与细胞表面受体结合,通过G蛋白介导的效应系统产生介质,活化丝氨酸/苏氨酸或酪氨酸激酶,从而传递信号。
根据底物蛋白质被磷酸化的氨基酸残基可将蛋白激酶分为:
根据是否有调节物参与蛋白激酶活性将蛋白激酶(PK)分为
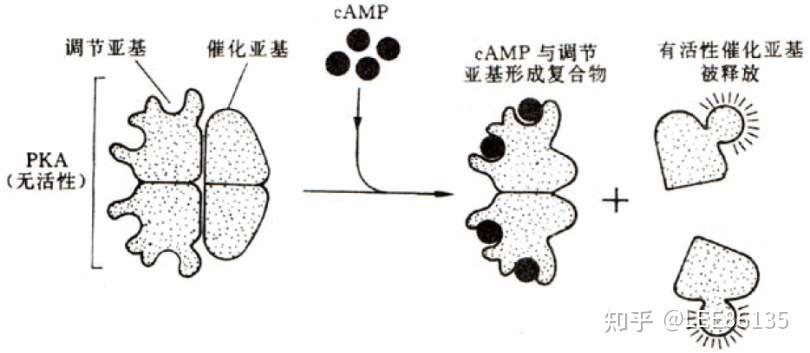
PKA
依赖于cAMP的蛋白激酶称为A激酶(PKA)。
A激酶能把ATP分子上的末端磷酸基团加到某个特定蛋白质的丝氨酸或苏氨酸残基上
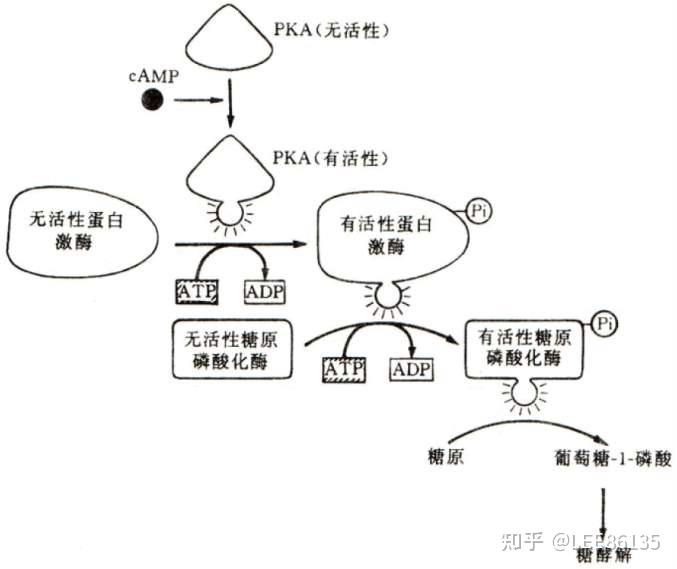
cAMP介导的蛋白质磷酸化过程
- 激素与其受体在肌肉细胞外表面相结合,诱发细胞质cAMP的合成并活化A激酶
- 活化的A激酶将活化磷酸基团传递给无活性的磷酸化酶激酶,活化糖原磷酸化酶
- 将糖原磷酸化,进入糖酵解过程并提供ATP
C激酶与PIP2、IP3、DAG
该蛋白激酶活性是依赖于$$Ca^{2+}$$的,故称C激酶(PKC)
磷酸肌醇级联放大的细胞内信使是PIP2(磷脂酰肌醇-4,5-二磷酸)的两个酶解产物:IP3(肌醇1,4,5-三磷酸)& DAG(二酰基甘油)
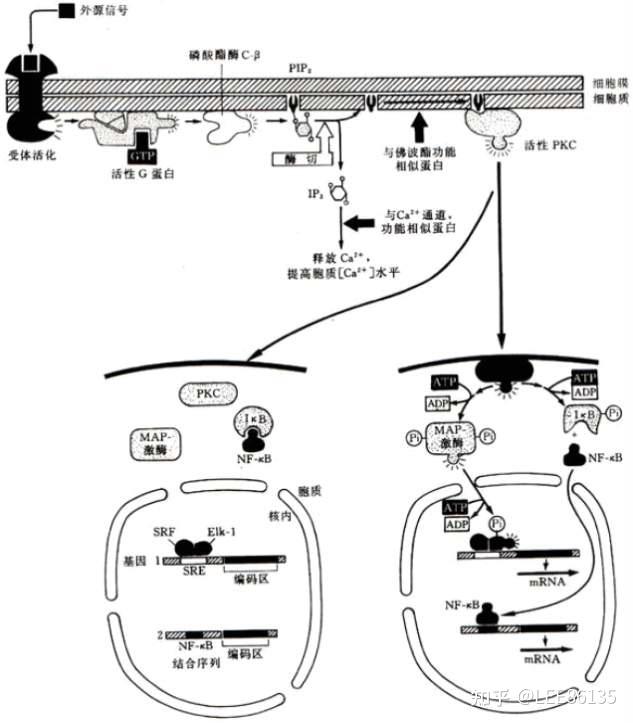
CaM激酶及MAP激酶
$$Ca^{2+}$$的细胞学功能主要通过钙调蛋白激酶(CaM)来实现的,它们也是一类丝氨酸/苏氨酸激酶, 但仅应答于细胞内$$Ca^{2+}$$水平。
MAP激酶(ERKS)活性受许多外源细胞生长、分化因子的诱导,取决于该蛋白中仅有一个氨基酸之隔的酪氨酸、丝氨酸残基是否都被磷酸化
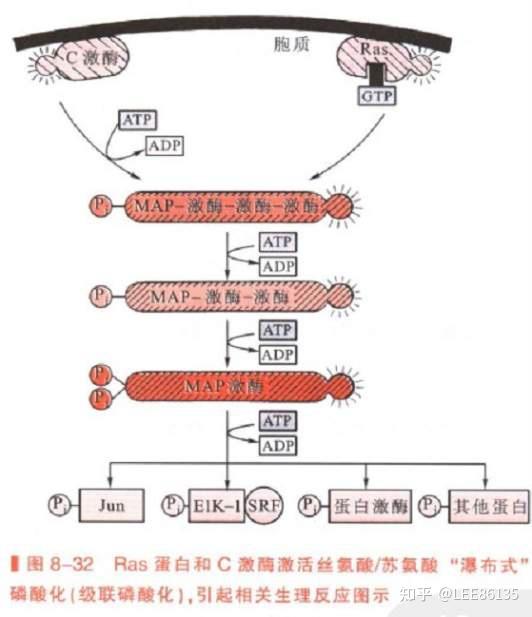
蛋白质磷酸化与细胞分裂调控
细胞通过p53及p21蛋白控制CDK(周期蛋白依赖的蛋白激酶 )活性,调控细胞分裂的进程
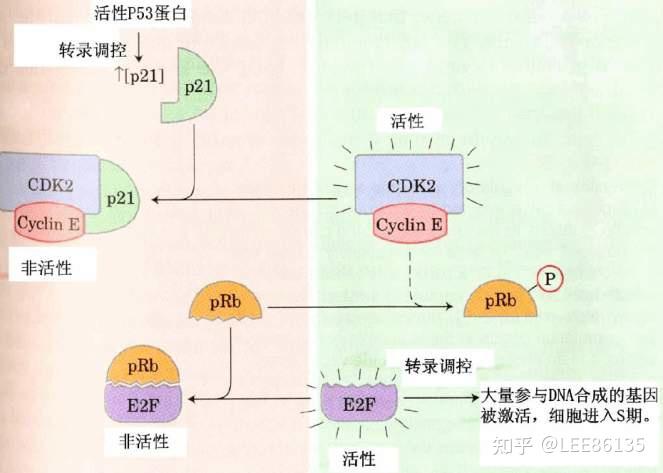
负:细胞中p53基因活性升高→p21蛋白过量→周期蛋白E-CDK2复合物不能将PRb蛋白磷酸化→PRb蛋白能与E2F相结合→细胞不能由G1期进入S期,细胞分裂受阻。
正:细胞中p53基因活性降低→p21蛋白下降→周期蛋白E-CDK2复合物能将PRb蛋白磷酸化→PRb蛋白不能与E2F相结合→细胞能由G1期进入S期,引发细胞分裂。
CDK(周期蛋白依赖的蛋白激酶)活性受周期蛋白和磷酸化双重调控
酪氨酸激酶(PTK)途径
- 跨膜受体家族--受体类由胞外结合配体结构域、跨膜结构域和细胞质激酶结构域组成,其PTK活性受胞外结构域与配体的调节。
- 胞质非受体家族--非受体酪氨酸激酶除有一段与前者同源的激酶结构域序列外,还有数个前者所不具备的保守区(Src同源结构域)
保守区Src亚家族非受体酪氨酸激酶及PTK作用底物分子都存在被称为Src同源结构域(SH)的序列,激酶功能区称为SH1,另外两个区分别命名为SH2和SH3。含有SH2结构域的蛋白也常常含有SH3结构域。
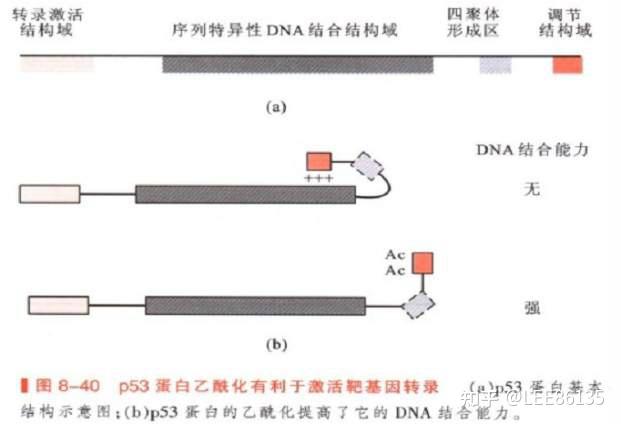
蛋白质乙酰化对转录活性的影响
乙酰化使p53蛋白(肿瘤抑制因子)的DNA结合区域暴露,增强了DNA结合能力,从而促进了靶基因的转录。
DNA甲基化诱导组蛋白去乙酰化,从而抑制靶基因转录
激素
许多类固醇激素以及一般代谢性激素的调控作用都是通过起始基因转录而实现的。
体内存在的许多糖皮质类激素应答基因都有一段大约20bp的顺式作用元件(激素应答元件,简称HRE),该序列具有类似增强子的作用,其活性受激素制约。
固醇类激素的受体蛋白分子有相同的结构框架,包括保守性极高并位于分子中央的DNA结合区,位于C端的激素结合区和保守性较低的N端。
激素、受体与顺式元件的结合位点三者缺一不可,其中无论是受体蛋白与激素的结合,还是激素本身,都不是与DNA结合并激活转录所必需的。
通常情况下,受体蛋白中激素结合结构域妨碍了DNA结合区及转录调控区发挥生理功能,只有与相应激素结合后才能打破这种障碍。
RNA的加工成熟
rRNA加工有两块:一个是分子内的切割,另一个是化学修饰。
真核生物rRNA基因转录时先产生一个45S的前体rRNA,然后前体rRNA被加工降解,生成不同相对分子质量的成熟rRNA。rRNA的化学修饰主要是核糖甲基化。
tRNA基因转录时也先生成前体tRNA,tRNA基因的初级转录产物在进入细胞质后,首先经过核苷的修饰,生成4.5S前体tRNA,再剪接成为成熟tRNA(4S)。
编码蛋白质的基因转录时首先生成前体mRNA,然后再加工剪接为成熟mRNA。
加工主要包括在mRNA的5&#39;末端加&#34;帽子&#34;,在其3&#39;末端加上poly(A),进行RNA的剪接以及核苷酸的甲基化修饰等
真核生物的基因可以按其转录方式分为简单转录单位与复杂转录单位。这两种转录方式虽然最终都产生蛋白质,但它们的转录后加工方式是不同的。
简单转录单位只编码产生一个多肽,其原始转录产物有时需要加工,有时则不需要加工
- 没有内含子,不需要转录后加工,其mRNA3’末端没有poly(A),但有一个保守的回文序列作为转录终止信号。
- 没有内含子,所编码的mRNA不需要剪接,但需要加poly(A)。
- 有内含子,需要进行转录后加工剪接,还要加poly(A)
复杂转录单位--编码组织和发育特异性蛋白质的基因除了含有数量不等的内含子以外,其原始转录产物能通过多种不同方式加工成两个以上的mRNA。
- 利用多个5’端转录起始位点或剪接位点产生不同的蛋白质
- 利用多个加poly(A)位点和不同的剪接方式产生不同的蛋白质
- 虽无剪接,但有多个转录起始位点或加poly A位点的基因
真核生物能否长时间、及时地利用成熟的mRNA分子翻译出蛋白质以供生长、发育的需要,是mRNA的有效性,这与mRNA的稳定性以及屏蔽状态的解除密切相关的。
翻译水平调控
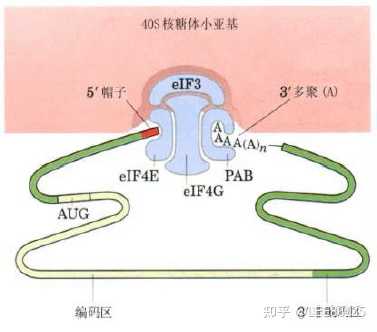
真核生物mRNA的“扫描模式”与蛋白质合成的起始
AUG的前和后序列特征为A/G NNAUGG,这使得核糖体滑行到mRNA的第一个AUG能准确识别并起始翻译。
mRNA 5&#39;末端帽子结构的识别与蛋白质合成
大多数真核生物mRNA5&#39;末端都带有“帽子”结构,所以,核糖体起始蛋白质的合成,首先面临的问题是如何识别这顶“帽子”。
真核生物mRNA5&#39;末端可有3种不同的帽子,即0型、1型和2型,其主要差异在于帽子中碱基甲基化程度的不同。
mRNA的稳定性与基因表达调控
高等真核生物转运铁蛋白受体(TfR)、铁蛋白负责铁吸收、铁解毒。
这两个mRNA上存在相似的顺式作用元件,称为铁应答元件(IRE)。IRE与IRE结合蛋白(IRP)相互作用,控制这两个mRNA的翻译效率。
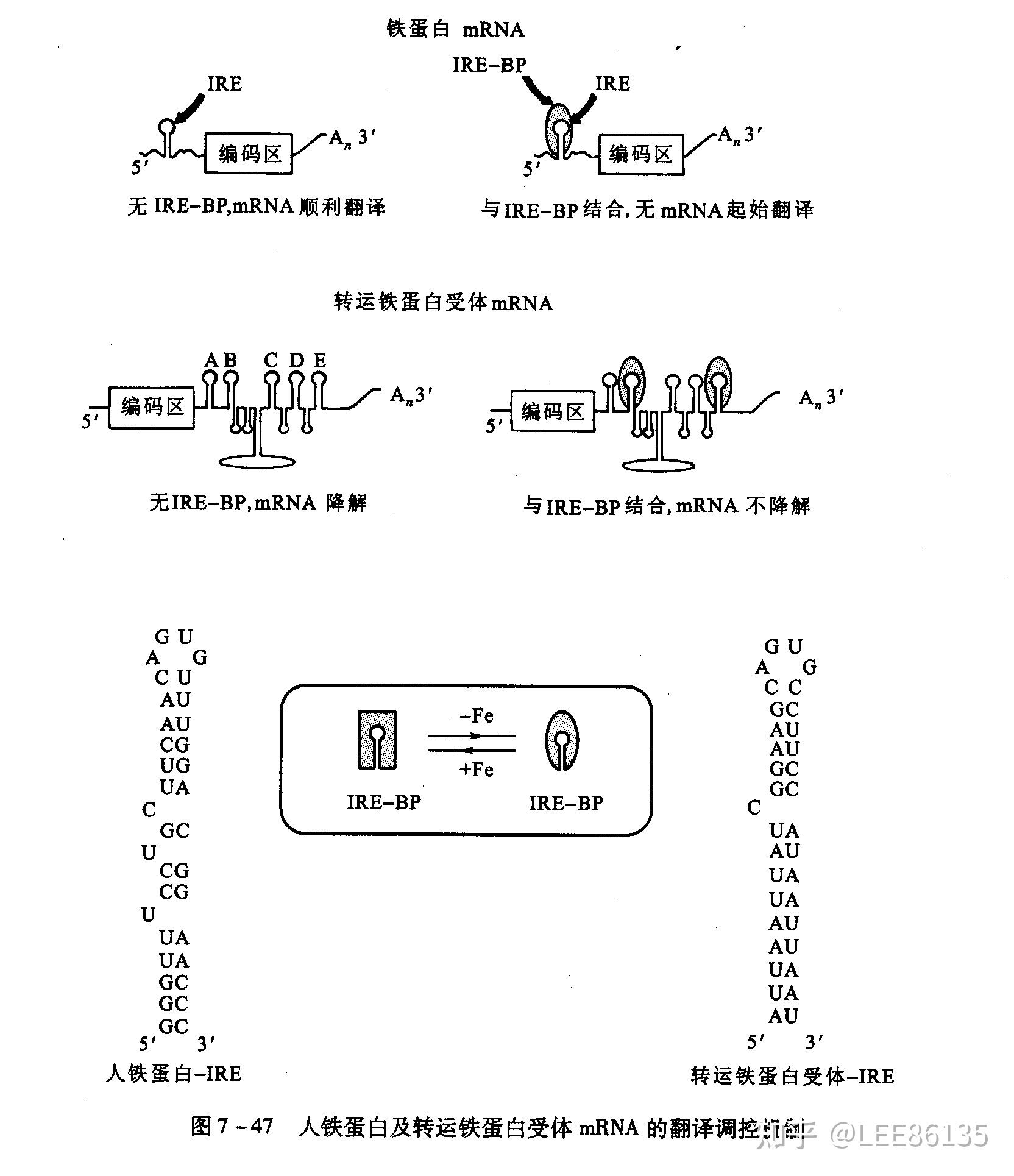
可溶性蛋白因子的修饰与翻译起始调控
许多可溶性蛋白因子(起始因子),对蛋白质合成的起始有着重要的作用,对这些因子的修饰会影响翻译起始。比如:
(1) eIF-2磷酸化对翻译起始的影响。
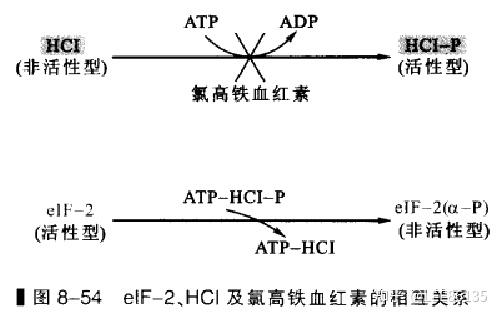
(2) CBPII活性与翻译的起始。
无法复制加载中的内容
蛋白质生物合成的调控,就是通过mRNA本身所固有的信号与可溶性蛋白因子或者与核糖体之间的相互作用而实现的。
第九章 疾病与人类健康 PPT已看
病毒癌基因(V-onc),编码病毒癌基因的主要有DNA病毒和RNA病毒(主要是反转录病毒)
细胞转化基因(原癌基因)(C-onc),它们能使正常细胞转化为肿瘤细胞,这类基因与病毒癌基因有显著的序列相似性
反转录病毒致癌基因
鸡肉瘤病毒(后称劳斯肉瘤病毒RSV)
反转录病毒颗粒结构
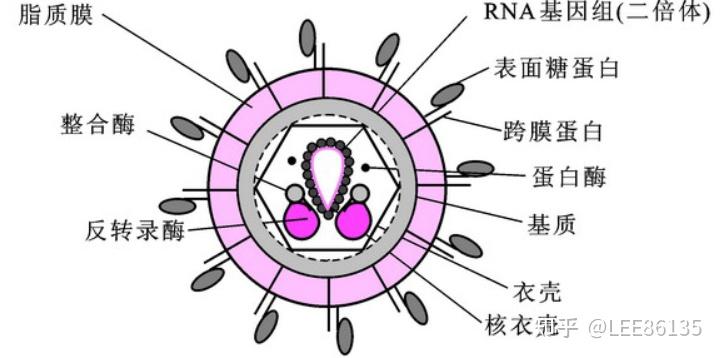
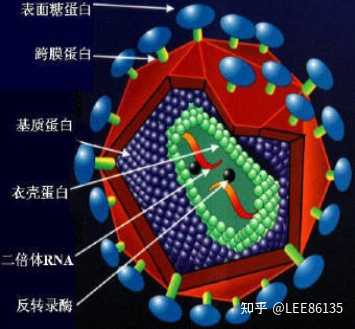
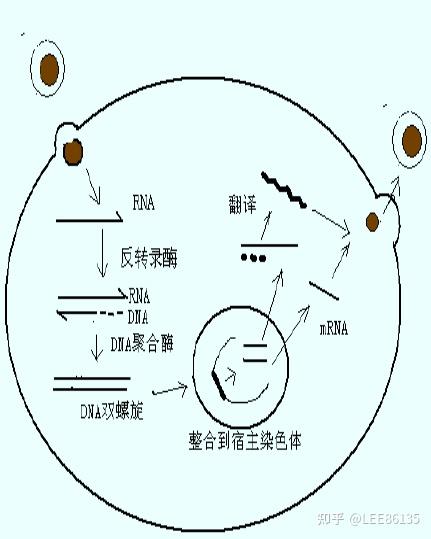
反转录酶以病毒RNA为模板,转录出单链DNA分子,利用宿主DNA聚合酶指导合成第二条DNA链,以双链DNA形式整合到宿主细胞基因组中
被整合的反转录病毒DNA分子称为原病毒,它能指导病毒mRNA的合成,并利用宿主细胞中的蛋白质合成机器,翻译生成病毒外壳蛋白等,最后组装成病毒颗粒
在肿瘤细胞中,“生长控制点”不起作用,所以瘤细胞一直处于细胞周期循环之中
原癌基因产物及其分类
细胞转化基因实质上就是由一类原癌基因突变而来
当原癌基因在某些外界因素(如放射线、化学致癌物等)的作用下,发生数量和结构上的微细变化,形成了致癌性的细胞转化基因,从而使正常细胞转化为肿瘤细胞
根据原癌基因产物在细胞中的位置,可将其分为3类:
原癌基因产物的一个共同特征:它们都能诱发一系列与细胞生长分化有关的基因表达,从而改变细胞的表型
表达调控
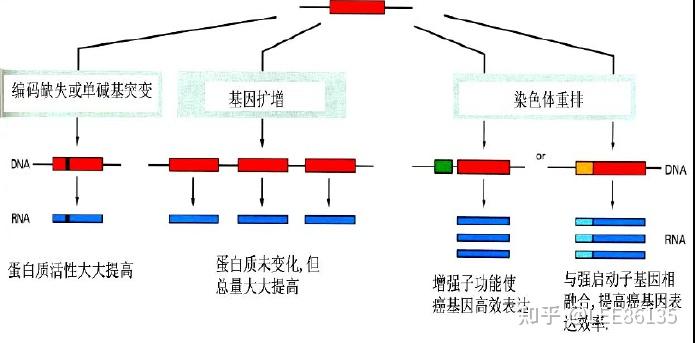
点突变
LTR插入
LTR是逆转录病毒基因组两端的长末端重复序列,含有强启动子序列,当LTR插入原癌基因启动子区域或邻近部位后,可从根本上改变基因的正常调控规律
基因重排
缺失
基因扩增
基因互相作用与癌基因表达
- 染色体构象对原癌基因表达的影响
- 原癌基因终产物对基因表达的影响
- 抑癌基因产物对原癌基因的调控
在功能上,抑癌基因起负调控作用,癌基因起正调控作用。在遗传方式上,原癌基因是显性的,抑癌基因是隐性的。在突变发生的细胞类型上,抑癌基因突变可以发生在体细胞中,也可能发生在生殖细胞中并通过生殖细胞得到遗传。原癌基因突变只发生在体细胞中。
HIV
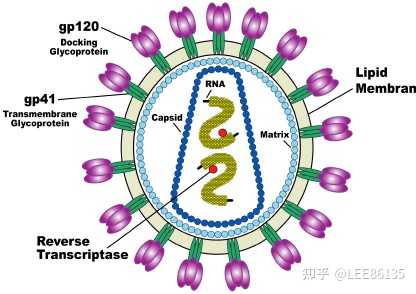
gp120和gp41为外膜蛋白,MA为内膜蛋白,CA为外壳蛋白,NC为核衣壳蛋白,RT为反转录酶
HIV基因为正链RNA,基因组9.7kb,5&#39;有帽子,3&#39;有polyA
主要功能区
复制
HIV主要浸染人体的T淋巴细胞及巨噬细胞,并通过病毒膜蛋白gp120与细胞表面的CD4受体蛋白结合
HIV导致免疫功能下降
- HIV颗粒的gp120脱落,与正常细胞表面CD4结合,被免疫系统误认为病毒感染细胞,导致正常细胞遭灭杀
- Gp120封闭T细胞CD4,影响T细胞免疫。
- Gp120刺激系统产生CD4特异性结合抗体,阻断T细胞功能
- 病毒胞膜导致细胞融合形成多核细胞丧失功能
治疗和预防
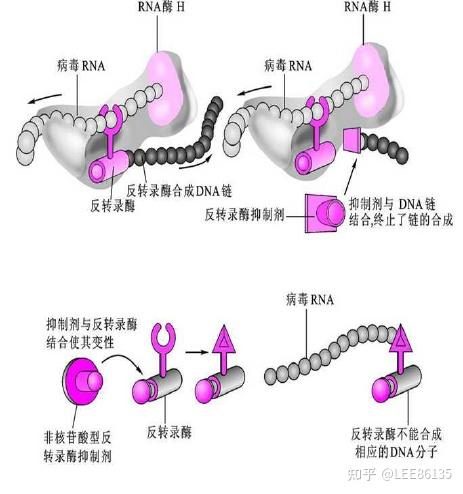
由于它在结构上与脱氧核苷酸的相似性,掺入后使病毒DNA的合成不能进行
与反转录酶相结合,通过限制该酶的移动性而影响它的活性
HBV乙型肝炎病毒
无法复制加载中的内容
HBV直径为42nm(Dane颗粒)由表面抗原、多糖和脂质构成。
根据血清反应(serologic reactions)的不同,分为adr、adw、 ayr、ayw 4种亚型
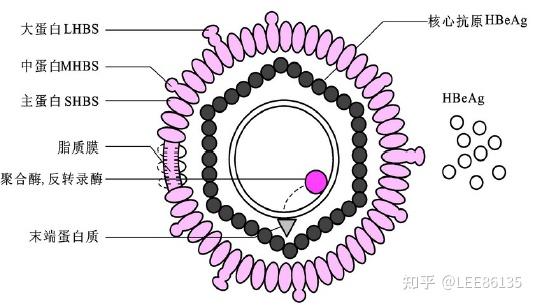
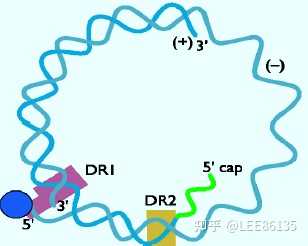
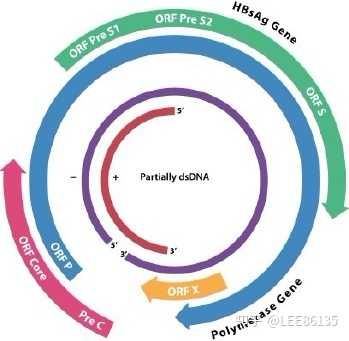
双链部分环状结构:两链长短不一
短链和长链的5′端通过240bp配对维持环状结构
长链(L)为负链,完整恒定,3.2Kb
短链(S)为正链,长度可变,约为长链长度的50~80%
两条链互补区两侧各有一个11个碱基的直接重复序列,称为DR1和DR2
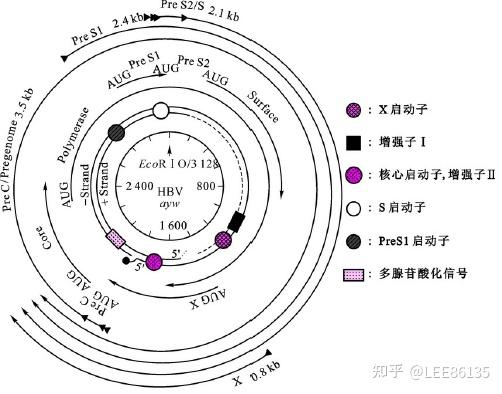
HBV在感染过程中分别产生3.5kb,2.4kb,2.1kb,0.8kb四种转录产物
HBV的编码区及产物
- S编码区。S编码区编码乙肝表面抗原蛋白。
- P编码区(最长编码区)。P编码区可能先翻译成多肽,然后加工成较小的功能型多肽
- C编码区。翻译产物为病毒核心抗原(HBcAg)
- X编码区(最小的编码区)。编码X蛋白
复制
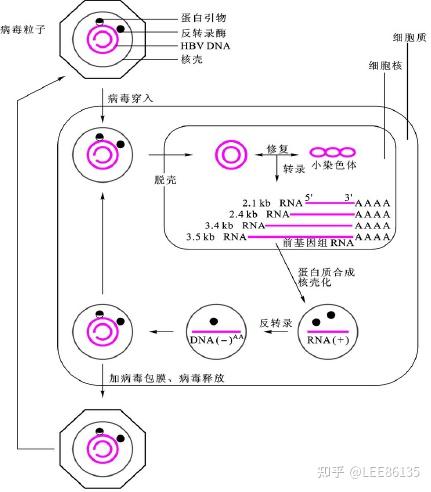
- 基因组DNA进入细胞核,形成闭环双链DNA并扩增
- DNA转录产生mRNA (RNA+),比DNA多130~270bp
- 包装,反转录形成cDNA(DNA-)
- 合成环状DNA
禽流感
正粘病毒分为3个属
- A型流感病毒:禽流感病毒,可对人、猪、马和禽致病
- B型流感病毒:仅能对人致病
- C型流感病毒:仅能对人致病
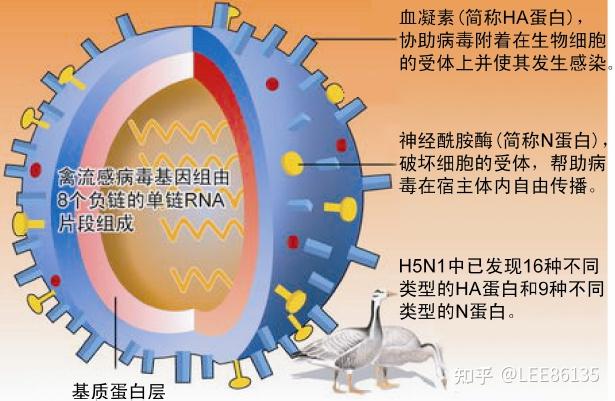
多呈球形,直径80~120nm
外层为包膜,中层为膜蛋白,内层为核心
单股负链RNA
禽流感病毒外膜表面有三种糖蛋白--血凝素(HA) 、神经酰胺酶(NA)、基质蛋白M2
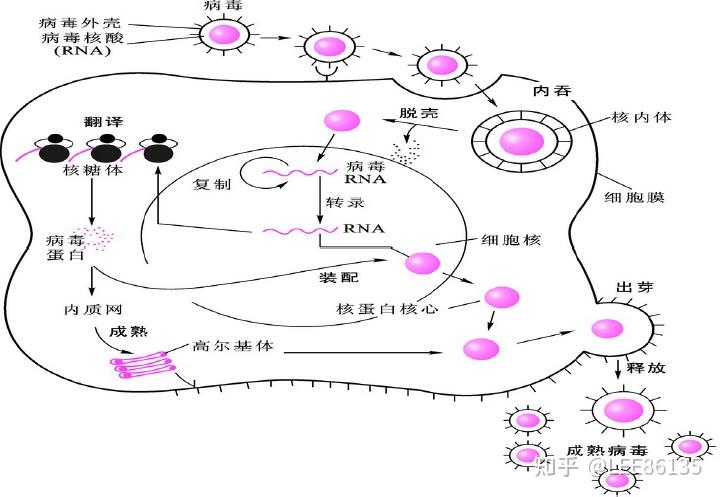
禽流感病毒进入细胞及转录与复制
- 病毒通过血凝素结合宿主细胞表面,进入细胞
- 进行复制
- 在宿主细胞内,新合成的病毒在细胞表面出芽
- 在神经酰胺酶作用下,从细胞释放
- 进行新的周期
禽流感病毒感染人类的机制
- HA受体结合位点的突变导致禽流感病毒易于感染人类细胞
- PB2蛋白627位氨基酸的点突变导致人禽流感病毒复制能力增强
- NSI第92位氨基酸的突变导致人禽流感病毒致病能力显著增强
严重急性呼吸系统综合症(SARS)
由SARS冠状病毒(SARS-CoV)引起。 SARS-CoV属冠状病毒科,冠状病毒属。SARS-CoV是一种单股正链RNA病毒,基因组全长29206个到29736个核苷酸。也是目前已知最大的RNA病毒。
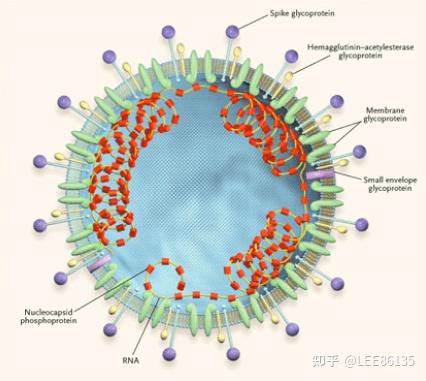
结构
S蛋白:刺突糖蛋白
E蛋白:小包膜糖蛋白
M蛋白:膜糖蛋白
N蛋白:核衣壳蛋白
HE:血凝素酯酶
RNA
无法复制加载中的内容
SARS-CoV的基因组结构
冠状病毒的核酸为非节段单链(正链)RNA,长27-31kb,是RNA病毒中最长的RNA核酸链,具有正链RNA特有的重要结构特征:即RNA链5’端有甲基化“帽子”,3’端有PolyA “尾巴”结构
SARS-CoV的侵染过程
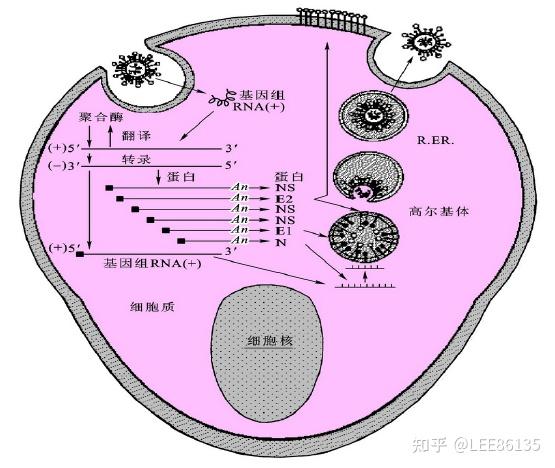
2019-nCoV
单股正链RNA病毒,入侵的细胞受体是ACE2
新型冠状病毒对免疫力和肺脏修复能力低的患者会造成严重损伤的原因--在肺脏,ACE2蛋白主要存在于Ⅱ型肺泡细胞和气道Club细胞这两种肺脏祖细胞表面,新型冠状病毒进入呼吸道后,通过S蛋白与ACE2结合,牢牢吸附并进入这些祖细胞内进行复制、增殖,并损伤肺脏再生系统。
检测
检测传染病的策略通常有两种:检测病原体本身,或检测人体为了抵抗病原体而产生的抗体。
检测病原体既可以检测抗原(一般是病原体表面蛋白,有些用内部核蛋白),也可以检测核酸。如果病人的体液中检测到抗体、抗原或核酸三者中的任何一种,则意味着已被感染。由于抗体产生需要时间,一般几天至几周不等,有些免疫功能较弱的病人可能抗体量很低,容易造成假阴性(已经感染,但是测试结果看起来没有被感染)。而在感染之初,病原体抗原含量也较低,同样不容易检测到。
目前广泛采用的是检测病原体的核酸序列,通过扩增反应来放大信号,检测的灵敏度高, 特异性好。
流行病学预测
病死率(Case fatality rate) --在一定时期内,因某种疾病而死的人数占确诊该疾病的人数的比例。
基本传染指数R(Basic reproduction number)--是病原体传染性强弱的一个指标,估算的是在传染病爆发初期(所有人都没有免疫的时候)一个有传染性的已感染者,平均可以直接传染多少人。
当R0<1时,传染病因为无法传播开而逐渐消失。
当R0=1时,传染病是地方性的,可控的,与人群长期存在
当R0>1时,传染病会迅速传播开,变得流行,如果不防控,就会指数增长
群体免疫(Herd immunity)--通常用在计算疫苗应当覆盖的人群比例上。$$R^*=R_0\times (1-H),R^*为人群接种疫苗之后的有效R_0,即拥有免疫的人群中,一个已感染者的平均传染人数$$$$H是人群中的免疫人群数量比例。当R^*=1时,临界值H=1-\frac{1}{R_0}$$
假设R0是2,那么至少需要50%的人群具有群体免疫。假设R0是4,那至少需要75%以上的人群具有免疫。
治疗方案
- 灭活疫苗
- 基因工程重组亚单位疫苗
- 腺病毒载体疫苗
- 减毒流感病毒载体疫苗
- 核酸疫苗
这次进入临床试验的疫苗,就是靠腺病毒载体疫苗研制的。
瑞德西韦(Remdesivir) 是核苷类似物,起初是吉利德公司开发用于埃博拉病毒的治疗,显示出一定的效果,但是总体疗效不如相关单克隆抗体,但安全性良好。在美国的个案报道中发现对新型冠状病毒有很好的作用,目前正在国内开展临床试验。
磷酸氯喹(Chloroquine) 于上个世纪40年代起用于治疗疟疾,后用于治疗类风湿性关节炎等。
磷酸氨喹在体外研究中已经展示出了非常好的抗新型冠状病毒活性。事实上,早些年一些研究表明,氯隆具有直接的抗病毒作用,抑制了包括黄病毒,逆转录病毒(如HIV)多种冠状病毒。在SRS流行时,不少研究发现氨喹具有潜在抑制ISARS病毒作用。
基因治疗
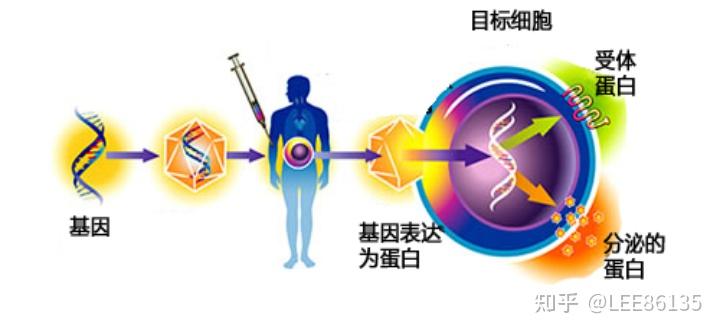
主要途径
将含外源基因的载体在体外导入人体自身或异体(异种)细胞,这种细胞被称为“基因工程化的细胞”,经体外细胞扩增后输回人体。
优点:容易操作,风险低。
缺点:不易形成大规模治疗。
将插入目的基因的表达载体直接导入体内
优点:简便省时省力,能很快观察到外源基因的表达及作用。
缺点:DNA在体内不易进入细胞,易被降解,以病毒颗粒的形式导入人体则易引起免疫反应和被补体灭活。
载体
基本条件
- 携带外源基因并能组装成病毒颗粒
- 介导外源基因的转移和表达
- 对机体没有致病能力
分类
I. 病毒载体:重组型病毒载体、无致病基因的病毒载体。
II. 非病毒载体
- 裸DNA
- 脂质体/DNA复合物
- 多聚物/DNA复合物
- DNA微球体
问题
基因治疗的关键问题是将治疗基因送入特定的靶细胞,并在该细胞中得到高效表达
导入基因后无调控表达将会产生严重后果。最理想的可控性是模拟人体内基因本身的调控形式
|
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2025-1-3 16:13
发表于 2025-1-3 16:13