DNA甲基化研究发展的时间线
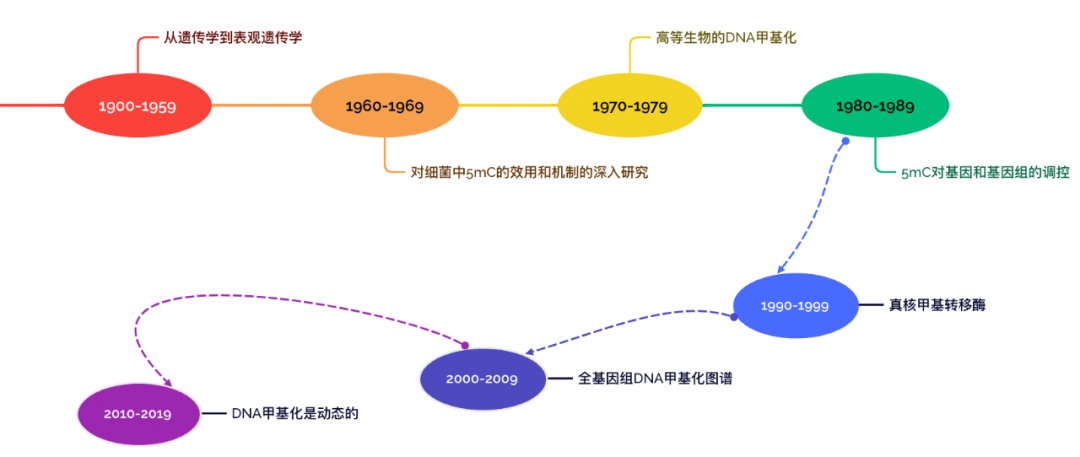
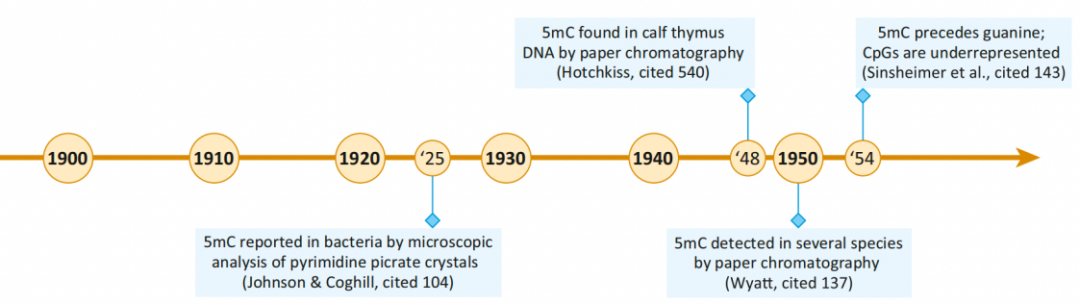
图1. DNA甲基化研究发展的时间线 DNA甲基化的发展历程 1900-1959:从遗传学到表观遗传学 1925年首次在结核分枝杆菌中发现5-甲基胞嘧啶,1948年通过纸层析法在小牛胸腺DNA中发现了5mC,两年后,证实了5mC在哺乳动物、昆虫和植物DNA中大量存在。1954年发现,5mC不是随机分布在DNA中,而是在CpG二核苷酸序列环境中特异发现的。
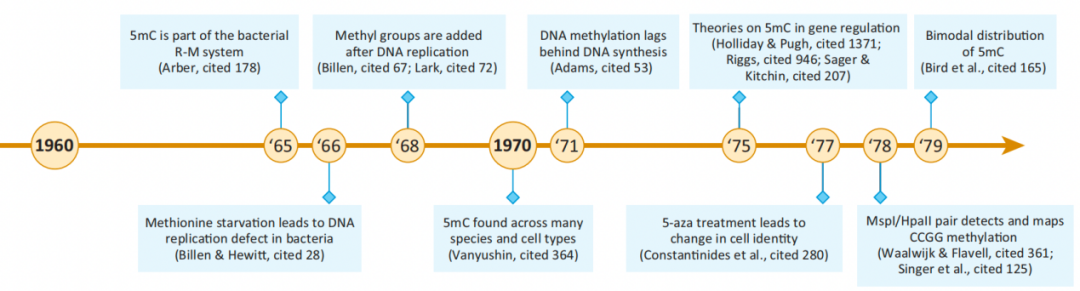
图2. 5mC发现的时间轴 1960-1979:对细菌中5mC的效用和机制的深入研究及高等生物的DNA甲基化 在此阶段发现了细菌的限制-修饰系统(R-M系统),其中甲基化敏感的“限制性内切酶”通过消化病毒的DNA来保护细菌宿主免受入侵病毒的侵害。除了在宿主保护中的作用外,还观察到细菌DNA甲基化与DNA复制之间的联系。 20世纪70年代,关于DNA甲基化在基因调控中的作用的研究和假设模型都取得了显著进展。改进的5mC检测方法实现了位点特异性甲基化分析,成为该领域的主要加速器。在本世纪末,该领域对物种内和物种间的DNA甲基化有了很好的认识,DNA甲基化作为基因表达的抑制因子被普遍接受。
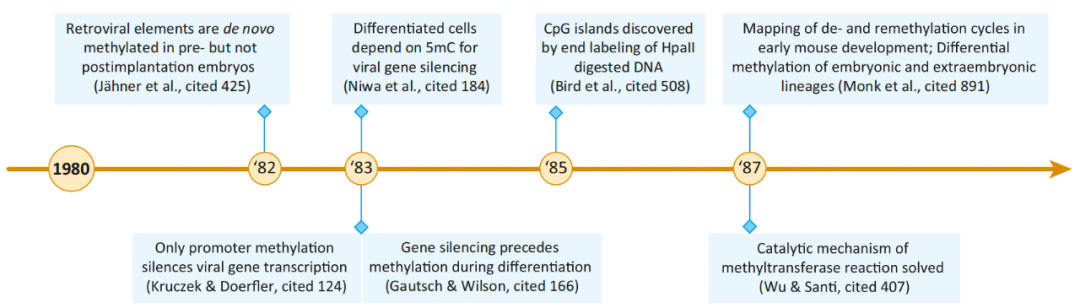
图3. 稳定甲基化模式的传播及其功能的初步理论 1980-1989: 5mC对基因和基因组的调控 20世纪80年代发现5mC的甲基化DNA的片段被非甲基化DNA的片段打断,每段片段占据不同的染色质部分。尽管精子总体上是高度甲基化的,但在精子细胞的组成表达基因中,可以看到像早期后生动物胚胎中发现的低甲基化延伸。随后的研究表明,这些未甲基化的区域由富含CpG的DNA组成,并且在许多小鼠组织中保持未甲基化,后来正式命名为CGI。1988年成功克隆出小鼠DNA甲基转移酶1 (Dnmt1),这是第一个哺乳动物DNA甲基转移酶。
图4. 启动子甲基化和CpG岛的发现 1990-1999:真核甲基转移酶 20世纪90年代,对基因组印迹和X染色体失活等现象的分子理解取得了重大进展,此外,一系列研究鉴定、克隆和敲除哺乳动物的甲基转移酶,揭示了它们在发育过程中的重要作用。1993年发表了DNA甲基转移酶M.HhaI的第一个晶体结构,同年科学家利用与小鼠和细菌甲基转移酶的序列同源性,鉴定并克隆了第一个植物DNA甲基转移酶MET1。 哺乳动物和植物DNA甲基转移酶和具有甲基结合结构域的5mC读取器的功能评估以及转录抑制的研究在20世纪90年代明确定义了。活跃的DNA是否以及如何发生甲基化,仍然没有明确的机制。
图5. 真核生物DNA甲基转移酶的发现和鉴定 2000-2009:全基因组DNA甲基化图谱 21世纪初,随着拟南芥、小鼠和人类基因组组装草图的完成,生物科学发生了重大转变。这些基因组图谱反过来又使研究表观基因组的其他层面成为可能,1992年首次报道的亚硫酸盐测序技术,成为量化和定位甲基化的金标准。2000年代中期在拟南芥中完成了第一个全基因组甲基化图谱,发现基因的5 '端大部分缺乏CpG甲基化,而组成表达基因的主体则广泛甲基化,CpG相对减少。 Bestor和Schübeler实验室使用类似的基于富集的技术创建了第一个人类DNA甲基化的综合图谱。Saxonov及其同事对人类基因组进行了计算分析,揭示了两类启动子:低CpG (LCG, 30%的启动子)和高CpG (HCG, 70%的启动子)密度启动子。与LCG启动子相关的基因往往具有独立于其启动子甲基化的转录潜力,并且大多数这些启动子通常被甲基化。相比之下,大多数HCG启动子被发现是未甲基化的。
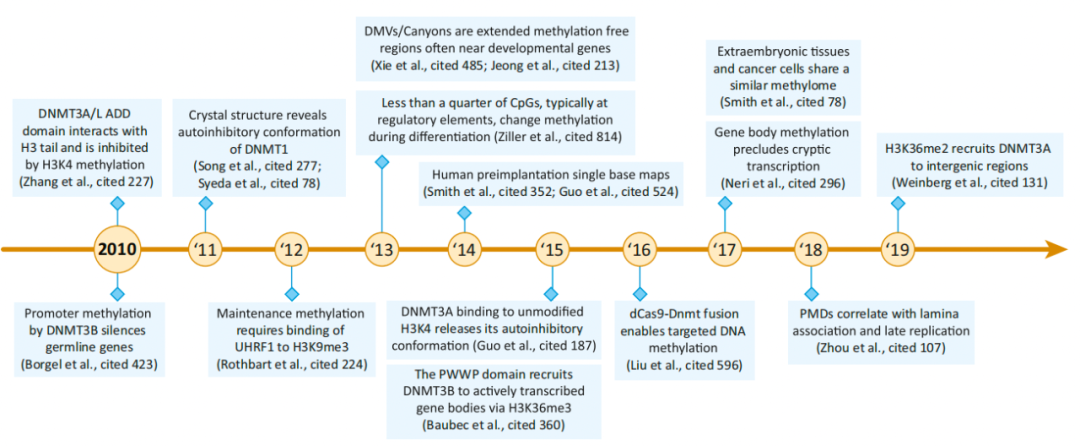
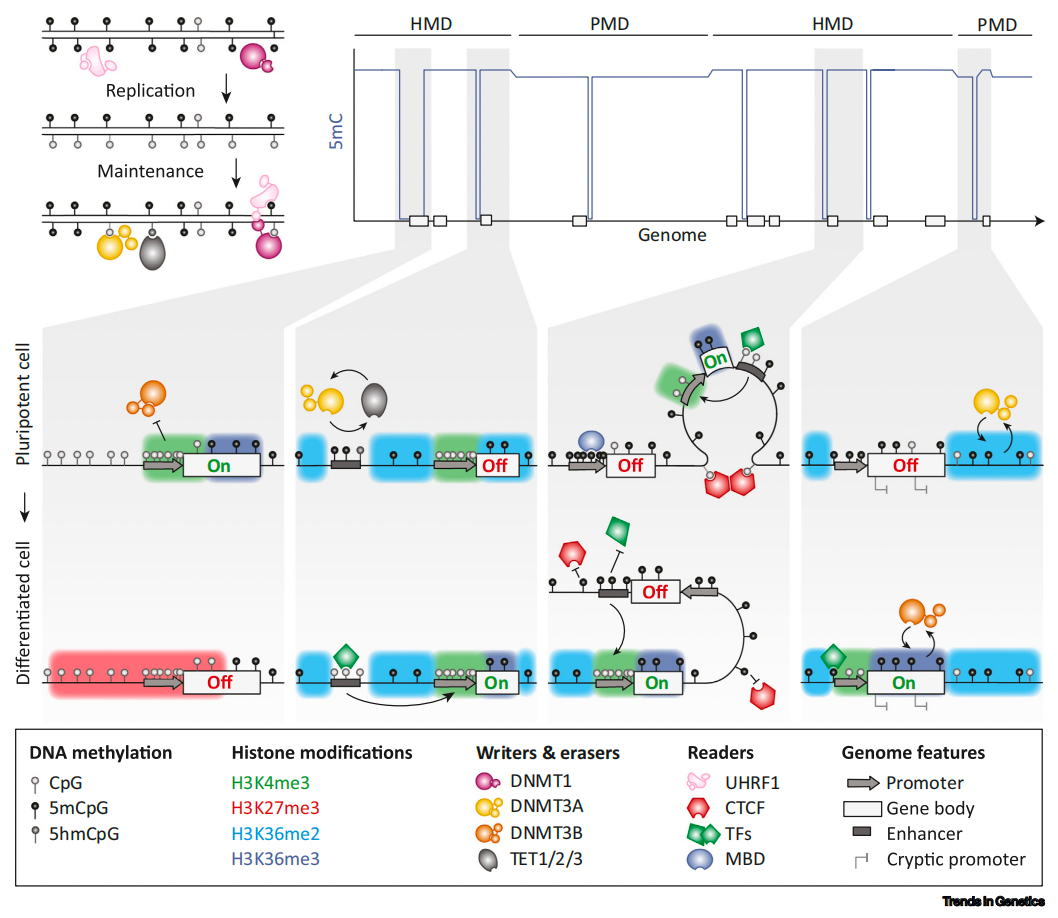
图6. 组蛋白修饰与5-甲基胞嘧啶(5mC)以及TET酶主动去甲基化的相互作用 2010-2019: DNA甲基化是动态的 测序技术的灵敏度、通量和可及性的提高使得一系列新的图谱研究成为可能,这些研究为许多新的见解提供了基础。陆续发现了DNA甲基化峡谷(dmv)、DNMT的变构调节、新的DNMT招募和靶点等。 更全面的5mC图谱改进了对高甲基化结构域(HMD)和部分甲基化结构域(PMD)的独特全局结构的描述。虽然PMDs可以在大多数健康细胞类型中检测到,但衰老、癌变和胚胎外组织都显示PMDs中甲基化水平进一步降低。结合结构研究,我们对DNMT的募集和活性以及DNA甲基化对基因调控的影响有了详细的了解。
图7. DNMT的招募机制和全局甲基化结构 结论 本综述提供了DNA甲基化领域从开始到现在的研究历程概述,并着重记录了该领域具有创新和巨大进步的关键实验,许多核心概念是在20世纪90年代建立起来的,最近的发现继续增加了新的见解(图8)。自从我们在细菌中发现5mC以来,对甲基化的研究持续了近一个世纪,DNA甲基化现在经常被用作研究其他生物过程和疾病表型的工具。尽管如此,仍有一些问题需要在未来解决。
图8. 哺乳动物DNA甲基化基因调控的关键概念 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号