前言 2025年6月28日,权威生物医学期刊《Life Medicine》在线发表了一篇由中国衰老标志物联合体(Aging Biomarker Consortium, ABC)撰写的专家共识文章——《A framework of biomarkers for visual system aging: a consensus statement by the Aging Biomarker Consortium》。该文章由中国多位视觉系统领域的顶尖专家共同完成,旨在系统性地梳理和总结视觉系统衰老的生物标志物,并为未来的研究和临床实践提供指导框架。 文章指出,视觉系统是人类感知世界的关键器官,随着年龄的增长,其功能会逐渐衰退,导至视力下降,严重影响老年人的生活质量。中国作为世界上人口老龄化最严重的国家之一,面临着巨大的视觉系统衰老相关疾病的挑战。为了应对这一问题,ABC联合体的专家们通过整合全球前沿研究和循证医学证据,结合临床经验,提出了一个涵盖功能、形态和分子三个维度的视觉系统衰老生物标志物评估框架。这一框架不仅有助于早期发现视力变化,还能为年龄相关眼病的进展提供重要线索,有望彻底改变老年社会视觉健康管理的现状,改善全球老年人群的健康结局。 文章详细介绍了视觉系统衰老的多维度特征,包括功能标志物(如睑板腺功能障碍、泪液功能障碍、老花眼等)、结构标志物(如角膜内皮细胞密度减少、晶状体变化、玻璃体液化等)和分子标志物(如糖化终末产物、视网膜Aβ沉积、基因多态性等)。此外,文章还提出了构建“视觉系统年龄”预测模型的思路,通过收集多维度数据,利用机器学习算法,为个体视觉衰老提供精准评估。这一创新性的研究方向将为未来的视觉系统衰老研究和临床应用开辟新的道路。
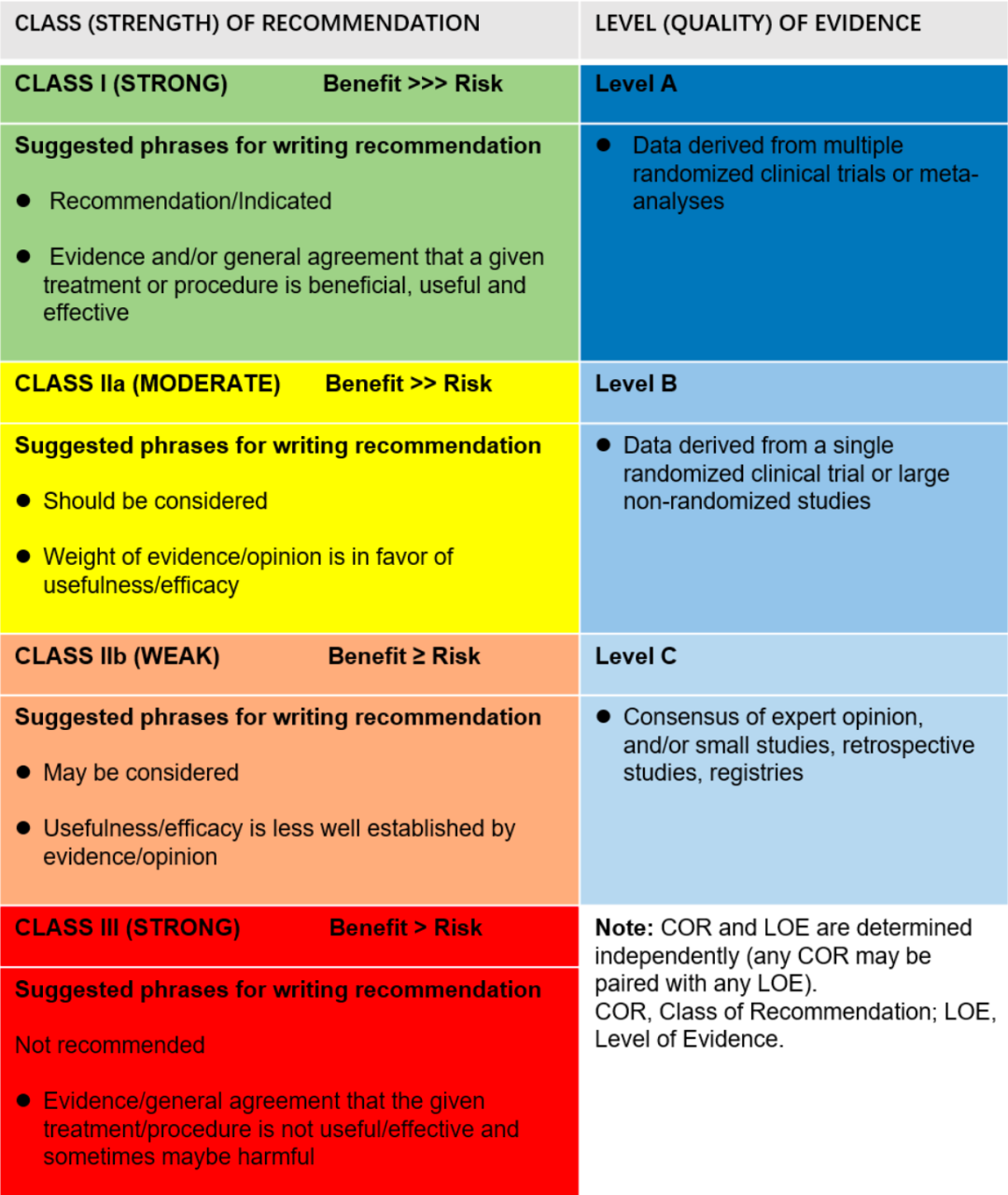
摘 要 视觉系统对人类感知至关重要,它将光信号转换为电脉冲并传输到大脑以处理环境信息。随着年龄的增长,其生理功能逐渐衰退,导至与年龄相关的视力障碍,严重影响老年人的生活质量。中国是世界上老年人口最多的国家,在通过有效治疗和干预对抗视觉系统衰老方面面临着巨大挑战。为应对这一挑战,中国衰老生物标志物联合体(ABC)整合全球前沿研究,综合循证医学与临床专业知识,制定了视觉系统衰老生物标志物专家共识。该共识提供了一个涵盖功能、形态和分子生物标志物的多维度评估框架。每个领域的经验证生物标志物不仅有助于早期发现视力变化,还能为年龄相关性眼病的进展提供见解。通过推进这一举措,ABC 旨在革新老龄化社会的视觉健康管理,最终改善中国乃至全球老年人的健康结局。 01 引言 视觉系统由生物体中专门的生理结构和神经机制组成,是感知和处理外部光信号的基本器官。视觉系统作为感光器官,将入射光子转化为电化学神经信号。这些信号通过视网膜 - 膝状体通路到达枕叶皮层,在那里被合成为连贯的视觉感知。然而,视觉系统的功能随着年龄的增长而逐渐下降,这涉及多个眼部结构的退行性变化。黄斑区的光感受器细胞和视网膜色素上皮(RPE)细胞功能下降,可能导至年龄相关性黄斑变性(AMD)等视网膜疾病的发生。此外,65 岁及以上人群中 AMD 的患病率超过 10%。此外,视网膜神经节细胞的进行性丢失和眼压调节减弱进一步增加了青光眼的风险 [1]。视觉系统的衰老过程与多种因素相关,包括认知能力下降、社会功能障碍和心理健康问题。这些情况对老年人的生活质量有显著影响 [2,3]。 作为世界上人口最多的老龄化社会,中国面临着与年龄相关疾病带来的前所未有的挑战。在现有的医疗框架中,缺乏标准化且能够准确量化视觉系统生理年龄、结构完整性和功能能力的生物标志物。目前针对与年龄相关的视觉系统疾病的临床干预措施有限。为解决这一问题,中国衰老生物标志物联合体(ABC)在北京组织了一次关于视觉系统衰老生物标志物的专家研讨会。专家小组通过整合全球前沿研究,综合循证医学与临床专业知识,制定了视觉系统衰老生物标志物共识声明。该共识提供了一个涵盖功能、形态和分子生物标志物的多维度评估框架。每个领域的经验证生物标志物被推荐用于评估:(1)当前衰老状态;(2)衰老速度;(3)疾病易感性。该共识声明还为未来的临床研究奠定了方法学基础。通过推进这一举措,ABC 旨在革新老龄化社会的视觉健康管理,最终改善中国乃至全球老年人的健康结局。 02 视觉系统衰老生物标志物的推荐方法 本共识中的推荐级别和证据级别按照国际公认标准呈现 [4]。证据级别和推荐强度的详细内容见表 1。所有建议都经过了 ABC 成员的严格审查和讨论,确保将不同的观点和考虑纳入本共识。 表 1. 推荐类别(强度)和证据级别
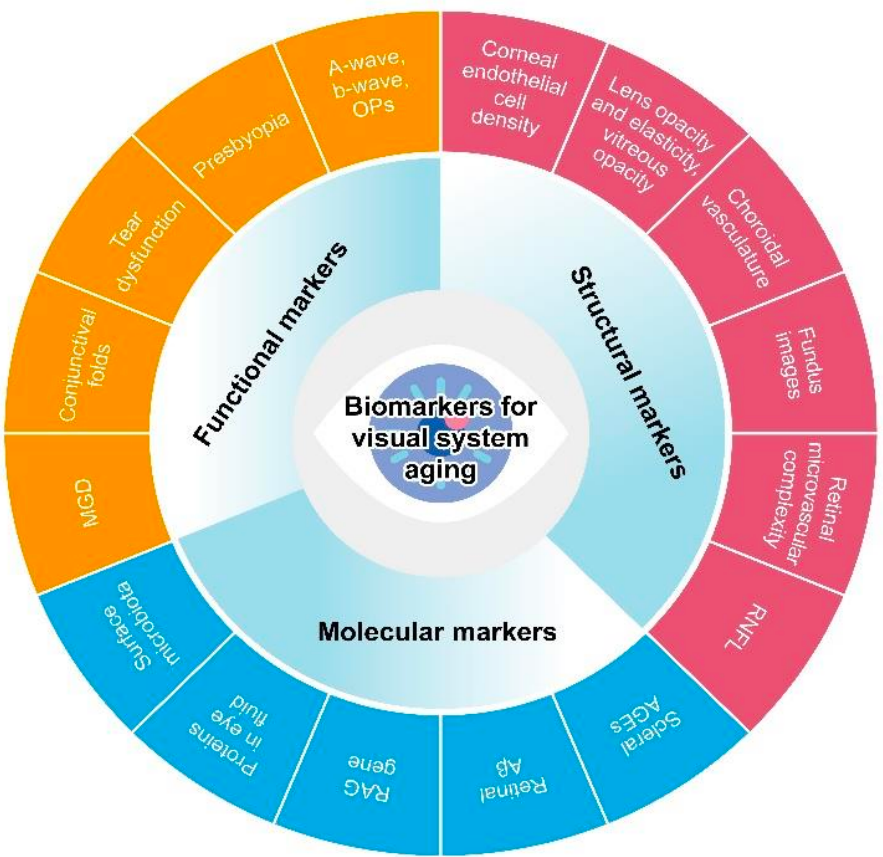
03 视觉系统衰老生物标志物的分类和临床应用 视觉系统的衰老是一个复杂的、多维度和多层次的生物学过程,涉及分子、细胞、器官和系统水平的改变。视觉系统衰老生物标志物作为关键的生物学指标,能有效反映视觉系统的实际年龄、功能和结构。这些生物标志物在预测视觉系统衰老的程度和速度、评估视觉系统相关疾病的风险以及评价抗衰老干预的疗效方面发挥着关键作用。本共识建立了一个综合评估框架,整合了功能、结构和分子生物标志物(图 1)。
图 1. 建议的视觉系统衰老标志物总结。建议的视觉系统衰老评估框架包括三个维度:分子、功能和结构生物标志物。它全面涵盖了视觉系统在衰老过程中各个层面发生的广泛变化。这些生物标志物有望在常规临床实践中得到广泛应用。然而,必须强调的是,需要额外的验证工作来评估这些生物标志物在视觉系统生物学衰老评估中的有效性。 3.1 视觉系统衰老的功能标志物 视觉系统衰老的主要临床表现包括视力下降、色觉异常和眼压升高。这些表现通常与各种视觉系统疾病的发生密切相关,特别是白内障、青光眼和黄斑变性,其患病率随年龄增长而上升 [5,6]。 3.1.1 眼睑退行性变化 衰老伴随着眼睑区域发生多种退行性变化,表现为上睑下垂、眼睑内翻或外翻、结膜松弛症和睑板腺功能障碍(MGD)等。最近的研究发现,衰老的睑板腺干细胞增殖信号下降,睑板腺神经支配不足,以及 niche 成纤维细胞中 I 型胶原蛋白的丢失 [7]。在人类和动物研究中均证实 MGD 的严重程度与衰老密切相关 [8]。根据《中国睑板腺功能障碍诊断与治疗专家共识(2023 年)》,中国 40 岁以上人群中睑板腺功能障碍的患病率为 54.7%-68.3%[9]。其评估基于多个参数,包括睑板腺开口、睑脂质量和睑板腺表达能力。人工智能(AI)可用于分析老年人睑板腺成像的形态参数,从而全面评估睑板腺状态 [10]。 结膜皱褶增多是另一种与年龄相关的退行性现象。随着年龄的增长,老年人常出现不同程度的结膜松弛,通常表现为结膜下组织松弛和折叠。一项日本流行病学研究显示,61 岁以上人群中结膜松弛症的患病率超过 98.0%[11]。同时,中国上海的一项研究发现,60 岁以上人群中结膜松弛症的患病率为 44.08%[12]。通过裂隙灯显微镜和眼前段光学相干断层扫描(AS-OCT)观察到,结膜松弛症的严重程度与年龄密切相关 [13]。临床上,通常采用睑缘平行结膜皱褶(Lipcof)分级系统来量化结膜皱褶的严重程度 [14]。 总之,MGD 和结膜皱褶增多可被视为视觉系统衰老的功能标志物。然而,必须排除非年龄相关的致病因素。 3.1.2 泪液功能障碍 随着年龄的增长,角膜神经密度逐渐降低,特别是在角膜上皮和基质层内。已证实这种下降会导至老年人的泪液功能障碍,这也可能与泪腺和睑板腺功能下降密切相关。流行病学研究表明,30-40 岁人群干眼症的患病率约为 20%,70 岁以上人群增至 36.1%[15]。如《中国干眼专家共识:定义与分类(2024 年)》所述,裂隙灯检查和泪膜破裂时间(TBUT)测试是评估干眼症状的重要方法(表 2)。因此,泪液功能障碍可能作为视觉系统衰老的一个潜在指标。全面的衰老评估应结合其他视觉系统退化标志物,以提供更全面的评估。 表 2. 中华医学会 2024 年干眼症分级
3.1.3 视觉功能异常 3.1.3.1 老视 睫状肌的调节功能以及晶状体在形状和硬度方面的稳定性,在维持清晰的近视力方面起着至关重要的作用。然而,随着年龄的增长,睫状肌的调节功能逐渐下降,主要表现为肌肉弹性降低、调节幅度减小和瞳孔反应减慢。此外,晶状体弹性的逐渐丧失加剧了眼调节能力的下降。 全球约有 18 亿人受到老视的影响,且这一数字仍在上升 [16]。老视通常在 45 岁左右开始出现,并随着年龄的增长而逐渐加重。大多数正视眼者在 45 岁时需要约 + 1.50D 的附加屈光力,50 岁时约为 + 2.00D,60 岁及以上时约为 + 3.00D。这种进展与衰老密切相关,表明它是随着自然衰老过程逐渐发生的生理现象。 睫状肌的功能通过测量眼睛从远点到近点的调节能力来评估。负相对调节(NRA)和正相对调节(PRA)可分别测试睫状肌的放松和收缩能力。同时,RAF 近点尺不仅可以检测晶状体调节幅度的下降,还可以监测调节近点的逐渐后移 [17]。 3.1.3.2 视网膜功能退化 黄斑区是眼睛负责精细中心视力的最关键区域,随着时间的推移容易发生退行性变化,导至中央视网膜功能丧失。临床上,这通常表现为视力模糊或中心暗点。视觉功能的综合评估包括视野检查以评估中心和周边视野缺损,以及视力、对比敏感度和色觉测试,以确保疾病诊断的准确性。 在细胞水平上,可使用视网膜电图(ERG)检测与年龄相关的视网膜变化,ERG 显示暗视 a 波和明视 b 波振幅逐渐降低,直接反映视杆和视锥光感受器细胞的功能下降 [18]。值得注意的是,振荡电位(OPs)作为视网膜内层完整性的关键指标,其变化先于 a 波和 b 波的改变,对与年龄相关的变化表现出更高的敏感性,特别是在低频波段。此外,Morlet 小波变换(MWT)作为一种先进的信号分析技术,是对传统时间 - 振幅分析的补充。这使其成为评估与年龄相关的视网膜功能下降的高度敏感工具 [19]。 推荐建议:
3.2 视觉系统衰老的结构标志物 视觉系统的衰老不仅表现为功能下降,还表现为组织结构的老化。这些结构改变往往是视觉系统疾病发生和发展的基础。因此,了解视觉系统衰老的结构特征对预测疾病风险具有重要意义。 3.2.1 角膜内皮细胞数量减少 随着年龄的增长,角膜的结构和功能会发生一系列退行性变化。主要特征包括角膜上皮和内皮层基底膜(特别是内皮层基底膜的后弹力层)增厚 [20]。此外,角膜上皮神经丛内的神经密度随年龄增长而降低。角膜内皮细胞数量减少及其形态异常都是老年人角膜衰老的重要特征 [21]。内皮细胞密度自然下降,从青年时期的约 3000-3500 个 /mm² 降至 60-70 岁时的约 2000 个 /mm²。此外,老年人角膜缘常出现新月形或环形灰白色混浊,称为 “老年环”,是角膜老化的典型表现。角膜内皮细胞密度的变化是视觉系统衰老的关键结构标志物,可通过活体共聚焦显微镜(IVCM)和光学相干断层扫描(OCT)进行评估。 3.2.2 晶状体变化 年龄相关性白内障(ARC)是一种进行性退行性疾病,其特征是与年龄相关的晶状体混浊和颜色变化。这种病理过程通常在 50 岁以上人群中出现,其发病率逐渐上升。 年龄相关性白内障的临床表现主要包括晶状体混浊、色素沉积和透明度降低,最终可能导至视力障碍。为有效评估年龄相关性白内障的严重程度和范围,临床上广泛采用晶状体混浊分类系统 Ⅱ(LOCSⅡ)和 Emery-Little 白内障分级系统 [22,23]。LOCSⅡ 分级系统根据晶状体混浊的程度和位置对白内障进行分类,而 Emery-Little 分类则通过评估晶状体核的颜色变化和硬度特征进行分类。此外,主观评分系统和眼前段成像分析系统也用于提供全面诊断。 3.2.3 玻璃体液化 研究表明,14-18 岁人群中约 20% 的玻璃体腔发生液化,80-90 岁人群中超过 50% 的玻璃体发生液化 [24,25]。这种与年龄相关的玻璃体液化可能导至玻璃体后脱离(PVD),最常见于中老年人群(45-65 岁)。严重的液化可能增加视网膜裂孔和牵拉性视网膜脱离的风险。重要的是,玻璃体混浊的逐渐积累往往会导至视觉模糊。 3.2.4 脉络膜血管变化 人类脉络膜血管会发生与年龄相关的改变,表现为脉络膜总体厚度、血管直径和脉络膜血流量的进行性减少。眼底检查通常显示红色反射明显变暗。同时,脉络膜毛细血管的血管密度显著降低,这也与趋化因子和补体基因的表达上调、巨噬细胞浸润增加以及巨噬细胞极化改变有关 [26]。这些脉络膜血管变化可靠地反映了视觉系统的衰老,为衰老评估提供了经验证的结构标志物。 3.2.5 视网膜的衰老变化 3.2.5.1 组织结构变化 已观察到视网膜的结构改变,包括特定神经元群体数量的减少,如内核层(IPL)、内丛状层(INL)、外丛状层(OPL)和外核层(ONL)。这种现象伴随着功能退化,并已成为退行性视网膜病变研究的关键指标。随着衰老进程的推进,视网膜逐渐变薄,特别是外核层的萎缩尤为明显,同时视网膜神经节细胞(RGCs)数量减少。对衰老视网膜神经节细胞的电子显微镜观察显示,轴突体积增加,轴突内线粒体的平均直径增大,线粒体密度降低,这使得视网膜神经节细胞更容易发生退化和丢失。光感受器是最脆弱的神经元类型之一,在老年小鼠中数量显著减少 [27]。此外,其他研究也报告了类似的发现,并进一步发现内核层明显变薄 [28]。 3.2.5.2 眼底图像变化 先前的研究采用基于眼底图像特征的分析方法来评估与年龄相关的眼底变化,从而阐明了衰老眼底的几个特征。例如,光学相干断层扫描(OCT)显示视网膜神经纤维层(RNFL)显著变薄。半自动视网膜分析软件和共聚焦显微镜检测到动静脉血管直径变窄,以及血管壁厚度增加。总的来说,这些特征表明视网膜结构和功能随着年龄增长而逐渐退化 [17]。 一项涉及 11,052 名健康参与者的大规模研究成功开发了基于眼底图像的深度学习(DL)模型。通过收集 19,200 张以黄斑为中心的 45 度非散瞳非立体眼底图像,该模型能够准确预测视网膜年龄。研究团队引入了创新的 “视网膜年龄差距(RAG)” 概念,定义为模型预测的视网膜年龄与实际年龄之间的差异,作为衡量视网膜衰老的可靠指标。在这项研究中,视网膜年龄差距的绝对值被限制在 3.55 年范围内 [29]。此外,研究表明视网膜年龄可以作为评估脑细胞衰老的窗口。对 35,834 名无帕金森病(PD)病史的参与者的分析显示,视网膜年龄差距每增加 1 年,患帕金森病的风险就上升 10%[30]。 有趣的是,视网膜微血管复杂性有望成为一种新颖且极具前景的生物学年龄生物标志物。通过使用手持非散瞳眼底相机,研究人员发现视网膜微血管复杂性随年龄增长而显著下降。在患有与年龄相关疾病的个体中,这种下降速度是生理性衰老个体的两倍以上。这一重要发现表明,早期识别加速衰老的个体可以进行及时干预,从而有效延缓与年龄相关疾病的发生 [31]。 3.2.5.3 视神经的形态变化 视神经纤维的改变类似于在中枢神经系统其他区域观察到的与年龄相关的神经纤维变化。这些变化主要以髓鞘结构的显著改变为特征,其中髓鞘细胞质增厚是最突出的特征之一,并一直持续到髓鞘最终解体。此外,与年龄相关的视神经纤维变化包括髓鞘相关糖蛋白的显著减少、髓鞘囊泡的形成以及髓鞘的磷酸化。这些结构改变可能会影响视神经纤维传递视觉信息的速度 [32-35]。 在哺乳动物中,视神经衰老主要以视网膜神经节细胞(RGCs)轴突的明显变化和视盘周围视网膜神经纤维层(RNFL)的逐渐变薄为特征 [36]。这种变化可以通过光谱域光学相干断层扫描(SD-OCT)检测到。虽然视神经的形态变化被认为是评估视觉系统衰老的潜在指标,但目前的研究在很大程度上仍局限于动物模型,需要进一步的临床研究来验证其实际应用价值。 3.2.5.4 视网膜色素上皮(RPE)细胞的形态异常 在 51 岁以下的人群中,黄斑区的大多数视网膜色素上皮细胞保持多边形形状和一致的形态,59% 呈现六边形结构。在 80 岁以上的老年人中,黄斑区六边形细胞的比例降至 52%。视网膜色素上皮细胞的排列发生改变,其形态变得不规则,但总体细胞数量保持稳定 [37]。视网膜色素上皮细胞形态异常可能反映视网膜衰老;然而,由于缺乏高效且无创的临床检测方法,目前不建议使用。 推荐建议:
3.3 视觉系统衰老的分子标志物 3.3.1 巩膜分子标志物 晚期糖基化终产物(AGEs)是糖 / 代谢物与生物大分子通过非酶促糖基化反应形成的化合物。它们在衰老、糖尿病、氧化应激和慢性炎症过程中常见积累,导至广泛的组织损伤。角膜胶原和其他眼组织(如晶状体和布鲁赫膜)中的晚期糖基化终产物水平随年龄显著升高 [38]。无创研究强调了巩膜中与年龄相关的晚期糖基化终产物积累,尽管这一过程比布鲁赫膜沉积进展缓慢 [39]。巩膜是老年人群体中进行体内晚期糖基化终产物分析的一个有前景的目标,晚期糖基化终产物可作为视觉系统衰老的潜在分子生物标志物。 3.3.2 角膜分子标志物 与年龄相关的角膜上皮变化表现为通透性显著增加和屏障功能逐渐受损,这主要是由于整合素亚基的分布改变,特别是半桥粒中 a6 和 β4 亚基的不连续表达模式 [40]。 在老年人中,角膜内皮细胞的 DNA 修复能力急剧下降,同时氧化应激标志物 8 - 羟基 - 2'- 脱氧鸟苷(8-OHdG)水平升高,表明眼睛中氧化应激的累积。这些影响加速了角膜老化,导至角膜内皮细胞密度降低和角膜基质变薄 [41]。DNA 修复缺陷的小鼠模型表现出与老年人类角膜相似的结构。线粒体 DNA(mtDNA)特别容易发生突变。UV-A 诱导的损伤引发氧化应激,导至线粒体 DNA 损伤和特征性突变(如线粒体 DNA T414G 突变),最终导至角膜混浊和硬化 [42,43]。 干细胞标志物 p63 在大鼠角膜中的区域分布存在年龄依赖性差异:幼年:中央(+)/ 周边(-);老年:中央(-)/ 周边(+)[44]。 上述非特异性标志物目前缺乏有效且安全的临床检测方法,因此暂不推荐使用。 3.3.3 晶状体分子标志物α- 晶体蛋白作为晶状体中的主要结构蛋白,属于小热休克蛋白家族,在维持晶状体透明度方面发挥分子伴侣作用。研究表明,40-50 岁之间 α- 晶体蛋白发生显著的结构改变,其特征是可溶性游离形式蛋白减少和高分子量蛋白丢失,最终促进蛋白聚集体的形成 [45]。作为白内障形成的关键因素,这些聚集体广泛结合于晶状体纤维细胞膜,阻塞膜孔结构,阻碍抗氧化剂的运输。同时,由于衰老,晶状体逐渐变黄,这主要是由于谷胱甘肽 - 3 - 羟基犬尿氨酸葡萄糖苷的积累。这种荧光团在晶状体核中积累,并与晶状体蛋白发生交联反应。值得注意的是,这些聚集体的积累增加了光散射,进一步加剧了晶状体透明度的丧失 [46]。由于缺乏临床可行的无创检测方法,不建议使用晶状体分子生物标志物。 3.3.4 玻璃体分子标志物玻璃体经历一个渐进性液化过程,这与多种眼部疾病相关,包括玻璃体后脱离、视网膜裂孔、视网膜脱离等。光学相干断层扫描(OCT)显示玻璃体衰老过程中的特征性形态改变:黄斑区玻璃体皮质变薄;旁黄斑区玻璃体皮质逐渐增厚 [47]。 作为一种细胞成分稀少的细胞外基质,玻璃体主要由胶原蛋白和透明质酸组成。主要的分子见解包括:
这些分子之间的相互作用机制需要进一步研究,以阐明它们在玻璃体衰老和相关病理过程中的精确作用。 3.3.5 视网膜分子标志物 3.3.5.1 蛋白质标志物 β- 淀粉样蛋白(Aβ)作为玻璃膜疣的核心成分,是视网膜衰老的精确生物学指标。新的证据表明,它与年龄相关性黄斑变性(AMD)和阿尔茨海默病(AD)等神经退行性疾病均有关联 [50,51]。实验结果表明,外源性 β- 淀粉样蛋白会引发视网膜色素上皮(RPE)细胞的特征性衰老表型 [52]。一项值得注意的临床研究(n=82 名参与者,141 只眼)通过结合血浆生物标志物(Aβ42/40 比值、p-tau181、p-tau217)和视网膜 β- 淀粉样蛋白正电子发射断层扫描(PET)成像,实现了对临床前阿尔茨海默病诊断 > 90% 的特异性 [53]。虽然 β- 淀粉样蛋白沉积有望用于评估视觉系统衰老,但全面的鉴别诊断需要整合眼底成像特征和全身代谢标志物。 CD47 是一种跨膜蛋白,通过与 SIRPα 结合形成 “别吃我” 信号,在人类和小鼠中也显示出与年龄相关的表达下降。在年龄相关性黄斑变性中,CD47 的下调可能促进黑素体吞噬细胞的形成,并推动视网膜色素上皮细胞的退化 [54]。 衰老的视网膜色素上皮细胞是年龄相关性黄斑变性的核心病理因素。骨形态发生蛋白 4(BMP4)主要在视网膜色素上皮细胞中表达,通过 Smad 和 TAK1-p38 通路介导氧化应激诱导的视网膜色素上皮细胞衰老 [55]。此外,凋亡相关蛋白 3(Apr3)与氧化应激和衰老有关,在视网膜色素上皮细胞的复制性衰老和 premature senescence 过程中其表达被诱导。此外,其在 ARPE-19 细胞中的过表达会加速细胞衰老 [56]。因此,需要进一步研究以提供 BMP4 和 Apr3 在生理性衰老中特定表达模式的理论基础。 3.3.5.2 遗传标志物 3.3.5.2.1 全基因组关联研究(GWAS) 视网膜年龄差距(RAG)不仅是视网膜组织衰老的有效指标,也是生物学衰老的全身性生物标志物,凸显了其作为综合衰老评估指标的潜力。一项具有里程碑意义的研究整合了两个大型人群队列 —— 英国生物银行和苏格兰泰赛德糖尿病审计与研究遗传学(GoDARTS)的数据,采用多性状全基因组关联研究(MTAG)分析。这项研究确定了 13 个与视网膜年龄差距显著相关的独立单核苷酸多态性(SNPs),特别是 SH3YL1 和 OCA2 的突变,可能有助于衰老的进展 [57]。这一发现为视网膜年龄差距作为视觉系统衰老的分子生物标志物提供了强有力的遗传证据。 3.3.5.2.2 单细胞转录组学 通过使用单细胞 RNA 测序(scRNA-seq)和转座酶可及染色质高通量测序(ATAC-seq)技术对超过 119,520 个单细胞进行综合分析,研究人员构建了详细的老年人类和灵长类动物视网膜单细胞图谱 [58]。该研究包括: 确定 11 种主要的视网膜细胞类型:
主要发现包括:
这些发现为细胞类型特异性衰老模式提供了革命性的解析,并揭示了年龄相关性视网膜变性的潜在治疗靶点。 3.3.5.3 表观遗传标志物3.3.5.3.1 调控区域的 CpG 岛甲基化超长链脂肪酸延长酶 2(ELOVL2)基因编码一种参与长链多不饱和脂肪酸合成的酶,在多种组织中表达。ELOVL2 基因调控区域的甲基化已被确定为血清中衰老的有力证据,使其成为一种有前景的表观遗传生物标志物 [59]。随着年龄的增长,观察到 ELOVL2 启动子中 CpG 岛的年龄依赖性高甲基化,同时其蛋白质表达逐渐下降。玻璃体内注射去甲基化剂 5-Aza-dc 可以逆转 ELOVL2 启动子的高甲基化,从而恢复其表达水平 [60]。尽管体液中 ELOVL2 启动子的甲基化模式能有效反映全身性衰老趋势,但缺乏组织特异性。因此,目前不推荐将这种生物标志物用于临床。 3.3.5.3.2 组蛋白组蛋白是染色质的基本结构成分,在基因表达调控和染色质结构维持中发挥着不可替代的作用。在人类和动物模型中,组蛋白缺失被视为视网膜色素上皮(RPE)衰老的一个显著标志。在衰老的视网膜色素上皮中,研究人员发现:
现有研究还表明,组蛋白修饰的失调与神经退行性疾病(如阿尔茨海默病和帕金森病)的发生有关。因此,组蛋白的表观遗传研究在阐明细胞衰老机制方面具有至关重要的意义。 3.3.5.4 miRNA 相关标志物微小 RNA(miRNAs)是真核细胞中一类高度保守的内源性非编码小 RNA 分子(通常为 19-25 个核苷酸)。这些分子在调节各种生物学过程中至关重要,包括细胞增殖、分化、代谢稳态、发育控制和疾病发病机制。值得注意的是,许多 miRNA 与视觉系统的年龄相关性疾病有关,因为它们影响多种生物学过程 [62]。 3.3.5.4.1 let-7 家族let-7 miRNA 家族是高度保守的 miRNA,在眼组织中广泛表达。研究表明,let-7 家族参与神经退行性疾病,其中 let-7d 通过调节神经炎症反应和凋亡途径对疾病进展有特别贡献 [63]。let-7d 以及 let-7b 和 let-7c 的表达水平分别在衰老的视网膜和玻璃体中升高。研究还表明,let-7 水平升高会增强 Müller 神经胶质细胞中转录后透明质酸的产生,从而影响视觉系统及其相关疾病。 3.3.5.4.2 miR-33miR-33 调节视网膜色素上皮中的胆固醇外流途径,其随年龄增长的渐进性升高导至脂质稳态失调,并导至年龄相关性黄斑变性的发生。ATP 结合盒蛋白 A1(ABCA1)是视网膜色素上皮细胞中的一种膜转运蛋白,也负责维持细胞膜中的脂质平衡。特别值得注意的是,miR-33 直接靶向 ABCA1 的 3' 非翻译区(3' UTR),抑制其表达。与年龄相关的 miR-33 上调抑制 ABCA1 表达,随后损害胆固醇输出并加剧视网膜色素上皮细胞中的脂质积累 [64]。靶向抑制 miR-33 可能减轻视网膜色素上皮层中的病理性胆固醇沉积并减少免疫细胞浸润,潜在地减缓年龄相关性黄斑变性的进展。 3.3.5.4.3 miR-34amiR-34a 通过在循环系统和神经系统中的上调表达而被确立为一种衰老生物标志物。在小鼠大脑中,miR-34a 的上调与神经退行性病变相关,并参与 p53 和 Sirt1 信号网络。作为中枢神经系统(CNS)的延伸,视网膜也表现出类似的 miR-34a 动态变化,在 24 个月时稳步上升至峰值 [64]。需要进一步研究以确定这些发现与人类视网膜衰老的转化相关性。 3.3.5.4.4 miR-146 家族miR-146 家族由 miR-146a 和 miR-146b 组成,是一对具有同源序列的 miRNA,在视网膜和脉络膜衰老过程中发挥关键作用。miR-146a 在成纤维细胞、脐静脉内皮细胞和小梁网细胞中增加,而 miR-146a 和 miR-146b 在视网膜色素上皮细胞中的水平从 2 个月到 48 个月均升高。进一步的见解表明,miR-146a 模拟物的过表达抑制视网膜色素上皮细胞中炎症因子 IL-6 和 VEGF-A 的表达,这表明 miR-146a 可能通过负反馈效应调节衰老过程中的炎症反应 [65]。 3.3.5.4.5 性别特异性 miRNA 标志物新出现的证据揭示了 miRNA 表达谱中的性别二态性,这可能在身体衰老过程中对视网膜结构和功能产生不同影响。 老年小鼠中性别依赖性表达模式包括:男性特异性上调:miR-27a-3p、miR-27b-3p、miR-20a-5p;非性别特异性下调:miR-20b-5p [66]。 这些性别偏向的 miRNA 改变可能导至随年龄增长对视网膜疾病的易感性差异。 3.3.5.5 辅酶标志物辅酶是一组小的非蛋白质有机分子,它们与酶可逆地相互作用,促进生化反应的催化。随着人们年龄的增长,辅酶水平和功能的显著变化会影响衰老过程,包括核黄素(维生素 B2)、辅酶 Q10(CoQ10)和烟酰胺腺嘌呤二核苷酸(NAD⁺)。 核黄素转化后产生两种必需的辅酶:黄素腺嘌呤二核苷酸(FAD)和黄素单核苷酸(FMN),两者在细胞能量代谢中都是基础的。作为代谢最活跃的组织之一,视网膜需要大量的能量来维持视觉功能,因此始终需要高水平的 FAD 和 FMN。然而,随着年龄的增长,视网膜色素上皮和视网膜中的 FAD 和 FMN 浓度逐渐下降,这种变化可能与能量代谢受损有关 [18]。 据报道,老年视网膜中辅酶 Q10 浓度损失约 40%,同时抗氧化能力受损和 ATP 合成效率降低 [67]。补充辅酶 Q10 可能部分改善与年龄相关的损伤。然而,辅酶 Q10 的体内检测在技术上仍然具有挑战性,这对其临床应用构成了障碍。 NAD⁺参与视网膜线粒体氧化磷酸化,并促进 ATP 的产生,以满足光照下的高能量需求。随着年龄的增长,NAD⁺水平下降,这一过程与烟酰胺磷酸核糖转移酶(NAMPT)的表达降低并行 [68]。针对 NAMPT 和 NAD⁺生物合成途径的治疗方法在减轻视网膜色素上皮衰老方面显示出前景,进一步强调了 NAD⁺的关键作用。 3.3.5.6 磷脂标志物对多个途径的代谢中间体的综合分析表明,甘油磷脂代谢物(如磷脂酰胆碱)在预测视网膜衰老方面在敏感性、特异性和准确性方面具有独特优势 [69]。尽管它们有潜力成为新型生物标志物,但侵入性检测方法尚未可用。 3.3.6 泪腺标志物在老年小鼠中,组织病理学分析显示导管周围的上皮细胞显著减少,单核细胞浸润占腺体区域的 11.0%±6.5%。超微结构检查进一步揭示导管周围纤维化、腺泡细胞胞质中异常脂褐素积累以及以肿胀和嵴断裂为特征的线粒体损伤。在分子水平上,导管周围单核细胞和血管内皮细胞显示出氧化损伤标志物(8-OHdG、4-HNE、HEL)的上调以及衰老相关蛋白(如 p38 和 p16)的增强表达 [70]。免疫学研究表明,泪腺和淋巴结中表达 MHC-II 的 B 细胞水平升高 [71]。泪腺功能衰退伴随着乙酰胆碱释放和蛋白质分泌的减少。同时,副交感神经和交感神经的密度和分布均随年龄增长而减少 [72]。这些发现为泪腺衰老的分子机制提供了重要见解。 3.3.7 眼内液标志物眼内液(包括泪液、房水和玻璃体)中的分子标志物由于其相对可及性,为评估视觉系统生物学状况提供了宝贵的见解。 一项研究招募了年龄在 19 至 93 岁之间的志愿者(n=75)。检测到的与年龄相关的升高包括:炎症介质(IL-8、IL-6、RANTES)、组织重塑因子(MMP-1)、通过蛋白质芯片分析评估了 9 种显著的蛋白质标志物 [73]。 然而,鉴于候选蛋白质的范围相对有限,未来的研究应扩大到包括其他生物标志物候选物,旨在更全面地了解衰老过程中泪液成分的变化。 另一项研究利用来自房水和玻璃体的 120 份液体活检样本,开发了一种前所未有的方法,名为 “多种蛋白质起源追踪表达(TEMPO)”,该方法能够发现眼内液中的细胞起源,并促进视觉系统的精确年龄预测。通过整合蛋白质组学、单细胞转录组学和 AI 算法,该研究取得了几项重要发现:6313 种具有非线性衰老模式的房水蛋白质的表征、26 种核心衰老相关蛋白质标志物的鉴定、AI 驱动的眼部衰老关键细胞通路的阐明 [74]。 总而言之,整合眼内液的多组学数据有助于微创监测和测试。然而,需要广泛的验证来确保这些发现的可靠性,这将需要实施大规模的队列研究。 3.3.8 微生物群标志物睑脂、结膜囊和眼睑皮肤的微生物组成存在差异。一项采用 16S rRNA 基因测序的横断面研究(n=36)揭示了不同的微生物特征:
这些发现表明,眼表微生物群可能作为视觉系统衰老的实用生物标志物,并且可以通过无创拭子取样后的高通量测序进行检测 [75]。鉴于这项研究的样本量有限,需要进一步的临床研究来评估其实际效果。 推荐建议:
表 3. 推荐的视觉系统衰老生物标志物
04 视觉系统衰老生物标志物的临床应用视觉系统衰老生物标志物具有广泛的临床意义,包括疾病的早期诊断、个性化治疗方案制定和治疗效果评估。 (1)早期诊断和筛查:识别视觉系统衰老生物标志物能够早期识别眼部异常,甚至在明显症状出现之前。 视觉系统的衰老过程是一个多方面的生物学过程,其生物标志物的探索和应用具有重要意义。通过综合筛查和分析,整合功能、结构和分子生物标志物,可以开发出强大的工具来加强眼科实践,支持年龄相关性视觉系统疾病的早期诊断、个性化治疗和针对性衰老干预。随着生物标志物技术的不断发展,这些进步有望建立更精确的诊断标准,从而能够早期检测、预防和管理与年龄相关的视觉系统疾病。 05 “视觉系统年龄” 预测模型的构建5.1 数据收集5.1.1 样本选择(1)选择不同年龄、性别和生活环境的个体作为研究参与者,以确保样本的多样性和代表性。 5.1.2 数据收集(1)对视觉系统进行全面评估,包括视力评估、眼底检查和眼压测量等,以全面评估视觉系统健康状况。 5.2 模型构建5.2.1 建模方法选择(1)可采用多元线性回归、支持向量机和随机森林等机器学习算法。 5.2.2 变量选择(1)除视觉系统衰老标志物外,模型中还可整合年龄、性别、生活方式、紫外线暴露史和视觉系统疾病史等变量。 5.2.3 模型训练(1)利用收集的数据训练模型并优化其参数,以便准确预测视觉系统年龄。 5.3 模型评估和优化5.3.1 评估指标(1)常用的评估指标包括均方误差、平均绝对误差、决定系数等。这些指标用于衡量模型预测的准确性。 5.3.2 模型优化(1)根据评估结果,可以通过调整参数、纳入额外变量和改进建模方法来完善模型。 5.4 模型应用5.4.1 应用模型建立的视觉系统年龄预测模型可应用于临床研究,为视觉健康评估、疾病筛查和个性化眼保健策略提供科学依据。 5.4.2 验证模型(1)收集额外的样本数据对模型进行验证。(2)随着时间推移和新数据的积累,不断更新和完善模型,以确保其稳定性和泛化能力,同时最大限度地减少数据偏差导至的预测误差。 通过将视觉系统衰老生物标志物整合到年龄预测模型中,可以更精确地评估个体的视觉衰老状况,从而实现视力问题的早期发现和针对性干预,为个性化眼健康管理提供支持。 06 结论与未来展望根据专家讨论,在功能、结构和分子三个维度中共推荐了 16 项视觉系统生物标志物。这些标志物将在未来进一步在不同年龄组中进行验证(见表 3)。 中国视觉系统衰老生物标志物研究的工作路线图明确了以下关键目标:(1)建立一个约 1000 人的全国多中心老年队列,以识别和验证视觉系统衰老生物标志物,开发检测技术和方法,确定中国人群视觉系统衰老生物标志物的参考值,预测视觉系统衰老的 “转折点”,并明确干预的最佳时间窗口。(2)利用人工智能建立视觉系统衰老评估模型和视觉系统衰老相关疾病预测模型,充分发挥影像学在视觉系统诊断中的独特优势。(3)推进产学研用深度合作,促进科研成果的转化和应用。 通过整合多学科方法和技术创新,我们有望开发出更精确的视觉系统衰老评估框架,为年龄相关性眼病的早期预防和管理提供科学依据,最终为 “健康老龄化” 战略愿景做出贡献。 作者贡献金子兵、张清炯和薛天构思了本共识。所有作者均参与了共识声明工作组,并在起草或修订过程中对 manuscript 做出了贡献。Chao Ma, Boxin Geng, Yangqing Zhang 和 Shan Li起草了 manuscript。所有作者均阅读并批准了 manuscript 的最终版本。
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号